Проводящие пути головного и спинного мозга
Нервные клетки, вступая в контакты друг с другом, образуют цепи нейронов - проводники импульсов. По цепи нейронов импульс движется в строго определенном направлении, что обусловлено строением самих нейронов и синапсов (так называемая динамическая поляризация).
Цепи нейронов, выходящие за пределы центральной нервной системы и связывающие ее с периферией тела, получили название проводящих путей. Учитывая динамическую поляризацию цепей, понятно, что проводящие пути бывают двух видов: центростремительные (чувствительные или восходящие), несущие импульс от периферии тела к головному мозгу, и центробежные (двигательные или нисходящие), несущие возбуждение от головного мозга к исполнительному органу (мышце). Поскольку нервная система работает по принципу рефлекса (см. «Рефлекторную дугу»), то двигательные и чувствительные пути взаимосвязаны на уровне центральной нервной системы посредством цепей вставочных нейронов (ассоциативные и комиссуральные нервные волокна). Именно так образуются сложные рефлекторные дуги.
Восходящие проводящие пути.Восходящие, или чувствительные, проводящие пути в зависимости от строения и места расположения рецепторов, от которых они начинаются, делятся на четыре группы.
Экстероцептивные пути (от лат exterus - наружный) - несут импульсы от покровов тела - чувство боли, температуры, осязания и давления.
Проприоцептивные пути (от лат. proprius - собственный) - проводят импульсы от аппарата движения - мышц, сухожилий, суставных капсул, связок, неся информацию о положении частей тела, размахе движения, напряжении мышц и пр.
Интероцептивные пути (от лат. interns - внутренний) - проводят импульсы от внутренних органов и сосудов, где хемо-, баро-, термо- и механорецепторы воспринимают изменения химического состава, давления, температуры и других показателей лимфы, крови и внутренних органов.
Чувствительные пути специфической чувствительности, несущие импульсы от органов зрения, слуха, вкуса и обоняния. В принципе эти пути являются экстероцептивными со сложно устроенными рецепторными аппаратами.
Латеральный спинно-таламический путь - проводящий путь боли и температуры (рис. 254).
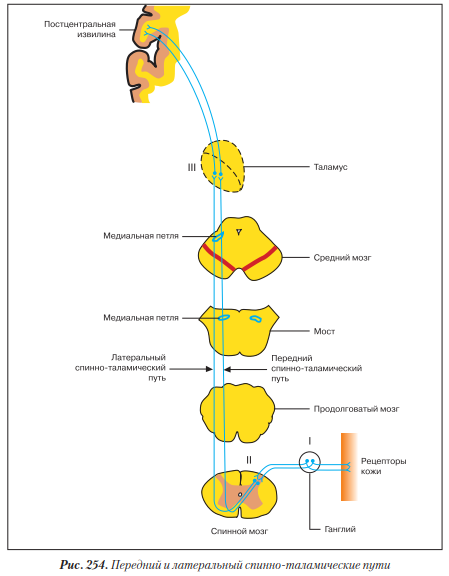
Первый нейрон - псевдоуниполярный, его тело лежит в ганглии. Периферический отросток (дендрит) заканчивается экстерорецептором боли или температуры в коже или слизистой оболочке. Центральный отросток (аксон) в составе заднего корешка спинного мозга заходит в серое вещество заднего рога, где переключается на II нейрон.
Тело второго нейрона лежит в собственном ядре заднего рога спинного мозга. Аксон в составе передней серой спайки переходит на противоположную сторону спинного мозга и поступает в боковой канатик, в составе которого поднимается в головной мозг. Проходит продолговатый мозг, мост и на уровне среднего мозга присоединяется к медиальной петле, в составе которой достигает таламуса, где переключается на III нейрон.
Тело третьего нейрона лежит в латеральном ядре таламуса. Аксон в составе таламокорковых пучков (пятый путь внутренней капсулы) проходит заднюю ножку внутренней капсулы и заканчивается в постцентральной извилине - ядре коркового анализатора общей чувствительности.
Поскольку латеральный спинно-таламический путь является полностью перекрещенным в спинном мозге, то при повреждении одной половины спинного мозга полностью исчезает болевая и температурная чувствительность на противоположной стороне тела.
Передний спинно-таламический путь - проводящий путь осязания и давления. Первый нейрон - псевдоуниполярный, его тело лежит в ганглии. Периферический отросток заканчивается экстерорецептором осязания или давления в коже и слизистой оболочке. Центральный отросток в составе заднего корешка спинного мозга заходит в серое вещество заднего рога, где переключается на II нейрон.
Тело второго нейрона лежит в собственном ядре заднего рога спинного мозга. Аксон в составе передней серой спайки переходит на противоположную сторону спинного мозга и поступает в передний канатик, в составе которого поднимается в головной мозг. Далее он проходит продолговатый мозг, мост, где присоединяется к волокнам медиальной петли, и в ее составе поднимается в средний мозг и достигает таламуса, где переключается на III нейрон.
Тело третьего нейрона лежит в латеральном ядре таламуса. Аксон в составе таламо- корковых пучков (пятый путь внутренней капсулы) проходит заднюю ножку внутренней капсулы и заканчивается в постцентральной извилине - ядре коркового анализатора общей чувствительности.
Особенностью переднего спинно-таламического пути в отличие от бокового является то, что часть аксонов вторых нейронов переходит на противоположную сторону не в спинном мозге, а в продолговатом. Другая часть аксонов вторых нейронов вообще не переходит на противоположную сторону, а идет в составе путей тонкого и клиновидного пучков (см. ниже). Поэтому при повреждении одной половины спинного мозга чувство осязания и давления на противоположной стороне тела полностью не исчезает.
Тонкий и клиновидный пучки - пути проприоцептивной чувствительности коркового направления (рис. 255).
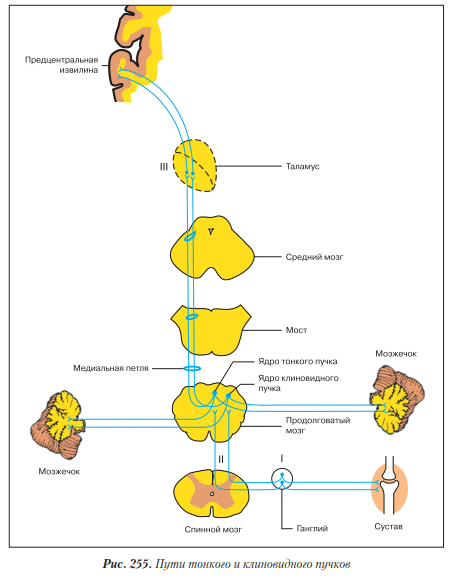
Первый нейрон - псевдоуниполярный, его тело лежит в ганглии. Периферический отросток заканчивается проприорецептором в аппарате движения - мышцах и суставах. Центральный отросток в составе заднего корешка спинного мозга заходит в белое вещество заднего канатика, по которому поднимается в продолговатый мозг к собственным ядрам тонкого и клиновидного бугорков, где переключается на II нейрон. Путь тонкого пучка начинается от нижних конечностей и нижней половины туловища, а путь клиновидного пучка несет импульсы от верхних конечностей и верхней половины туловища.
Тело второго нейрона лежит в собственных ядрах тонкого и клиновидного бугорков продолговатого мозга. Аксоны вторых нейронов формируют два вида дугообразных волокон.
1. Наружные дугообразные волокна переходят на противоположную сторону продолговатого мозга и формируют так называемую медиальную петлю, в составе которой проходят мост, средний мозг, и достигают таламуса, где переключаются на тело III нейрона. Место перекрещивания этих волокон получило название перекрест петель.
2. Внутренние дугообразные волокна идут по-разному (см. «Продолговатый мозг»):
Передние внутренние дугообразные волокна переходят на переднюю поверхность продолговатого мозга, затем на противоположную сторону, обходят спереди пирамидные пути, ядра олив и в составе нижней мозжечковой ножки достигают коры червя мозжечка.
Задние идут по своей стороне через нижнюю мозжечковую ножку к коре червя мозжечка своей стороны.
Тело третьего нейрона лежит в латеральном ядре таламуса. Аксон его в составе тала- мокорковых пучков (пятый путь внутренней капсулы) проходит заднюю ножку внутренней капсулы и заканчивается в пост- и предцентральной извилинах - ядре анализатора общей чувствительности и в ядре двигательного анализатора.
Таким образом, пути тонкого и клиновидного пучков проводят проприоцепторное чувство от аппарата движения в кору больших полушарий, что позволяет оценивать положение тела в пространстве. Наличие внутренних дугообразных волокон, которые заканчиваются в подкорковых центрах - коре мозжечка, есть проявление принципа множественного дублирования функций, свойственного нервной системе человека. Способность к движению - важнейшая функция живого организма, необходимое условие его существования. Поэтому пропри- оцепторные пути дублируют друг друга. С одной стороны, кора мозжечка связана с корой больших полушарий. С другой - внутренние дугообразные волокна дублируют спинно-мозжечковые пути, несущие бессознательное проприоцепторное чувство, что позволяет замкнуть рефлекс на уровне мозжечка. Так формируются подсознательные (автоматические) двигательные рефлексы, необходимые человеку как в повседневной жизни, так и на производстве или в спорте.
Передний и задний спинно-мозжечковые пути - пути проприоцептивной чувствительности мозжечкового направления (рис. 256).
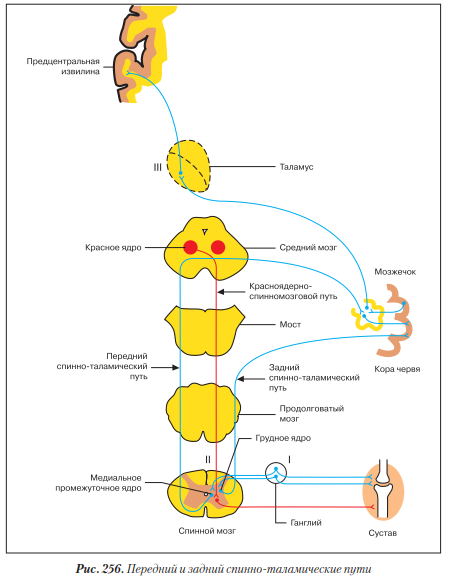
Первый нейрон - псевдоуниполярный, его тело лежит в ганглии. Периферический отросток заканчивается проприорецептором в аппарате движения - мышцах и суставах. Центральный отросток в составе заднего корешка спинного мозга заходит в серое вещество, где переключается на II нейрон.
Второй нейрон. Место расположения тела и ход аксона второго нейрона у переднего и заднего спинно-мозжечковых путей отличаются друг от друга.
Задний спинно-мозжечковый путь наиболее короткий. Тело второго нейрона лежит в грудном ядре задних рогов спинного мозга. Аксон выходит в боковой канатик и по задней его части поднимается к продолговатому мозгу, откуда по нижней мозжечковой ножке своей стороны достигает коры червя, где и заканчивается.
Передний спинно-мозжечковый путь более сложный, дважды перекрещенный. Тело второго нейрона лежит в медиальном промежуточном ядре бокового рога спинного мозга. Аксон в составе передней серой спайки переходит на противоположную сторону спинного мозга и затем по боковому канатику поднимается к продолговатому мозгу, проходит его, затем мост и на уровне среднего мозга опять переходит на противоположную сторону и уже в составе верхней мозжечковой ножки своей стороны достигает коры червя, где заканчивается.
По этим путям мозжечок получает информацию о состоянии опорно-двигательного аппарата, участвует в рефлекторной координации движений, обеспечивающей равновесие тела.
Ответная бессознательная двигательная реакция идет по мозжечково-красноядерно-спинномозговому пути. Импульс от коры червя по ассоциативным волокнам поступает в кору полушарий мозжечка, а оттуда - в зубчатое ядро мозжечка. От зубчатого ядра и от коры червя импульс поступает к красному ядру противоположной стороны. От красного ядра начинается красноядерно-спинномозговой путь, двигательный, экстрапирамидный, бессознательный (см. ниже).
Также можно проследить связь мозжечка с высшими подкорковыми и корковыми центрами проприоцепторной чувствительности. От зубчатого ядра аксоны через верхнюю ножку мозжечка поднимаются в средний мозг, затем переходят на противоположную сторону и заканчиваются в ядрах таламуса. Оттуда по уже описанным ранее таламо-корковым пучкам достигают коркового ядра проприоцепторной чувствительности (пост- и предцентральной извилины).
Дата добавления: 2022-01-27; просмотров: 1222;