Функциональная организация слуховых органов насекомых
Тимпанальные органы. У насекомых, относящихся к разным отрядам, в строении и функциональной организации тимпанальных органов имеются существенные различия, поэтому мы рассмотрим их в систематическом порядке.
Orthoptera Tettigoniidae. Исследование тимпанальных органов кузнечиков в значительной мере затрудняется тем, что к ним примыкают две большие группы хордотональных сенсилл подколенного органа, причем одна из них имеет с тимпанальным органом общую иннервацию (см. рис. 13). Аутрумом (Autrum, 1941) было установлено, что подколенный орган реагирует преимущественно на низкочастотные вибрации субстрата, а слуховой гребень (см. рис. 13) воспринимает звуковые колебания сравнительно высоких частот. Однако применявшиеся в этих экспериментах методы разрушения хордотональных сенсилл и отсутствие морфологического контроля лишили автора возможности четко разграничить функции двух групп рецепторов и выяснить функциональное значение промежуточного органа (см. рис. 13).
Исследования Кацуки и Суга (Каtsuki, Suga, 1960) показали, что у кузнечика Gampsocleis buer- geri в I грудной ганглий входят две группы слуховых афферентных волокон. Одна из них состоит из сравнительно крупных нейронов, обладающих оптимумом чувствительности на 10 кГц, а другая представлена тонкими волокнами с оптимумом чувствительности в диапазоне 6—7 кГц. Найденные клетки не были идентифицированы с известными группами сенсилл тимпанального органа, что дало основание для предположения о принадлежности этих нейронов подколенному и тимпанальному органам (Autrum, 1960).
Таким образом, после предварительных исследований оставалось неясным, участвует ли подколенный орган передних голеней в рецепции звука и существуют ли какие-нибудь функциональные различия между основными группами сенсилл в тимпанальном органе. Чтобы ответить на эти вопросы, мы провели серию электрофизиологических экспериментов, в процессе которых перерезали веточки тимпанального нерва (Жантиев, 1971).
В опытах были использованы кузнечики трех видов: Tettigonia cantans, Decticus verrucivoras, и Pholidoptera pustalipes, Подведя электрод под участок ножного нерва у места вхождения его в I грудной ганглий у Т. cantans, мы прежде всего зарегистрировали суммарную реакцию всех рецепторных элементов, отвечающих на звук. Затем, чтобы выяснить, какие рецепторы отвечают на звуковые стимулы, мы последовательно исключали из суммарной реакции импульсацию перечисленных ранее групп хордотональных сенсилл.
В первой серии экспериментов была проведена регистрация активности нейронов в тимпанальном нерве (рис. 19). Это позволило исключить из ответов возможную реакцию на звук проксимальной части подколенного органа. При сравнении полученных частотно-пороговых кривых с суммарными кривыми между ними не было ,найдено существенных различий.
Перерезка подколенной ветви тимпанального нерва (см. рис. 13) позволила полностью изолировать подколенный орган от тимпанального и наблюдать активность только двух групп сенсилл — промежуточного органа и слухового гребня (регистрация от тимпанального нерва). Полученные в этом случае ответы и частотно-пороговые кривые практически ничем не отличались от суммарных (т. е. полученных до операции), поэтому мы пришли к заключению, что подколенный орган не воспринимает звуковых колебаний в диапазоне 1—80 кГц интенсивностью до 90 дБ.
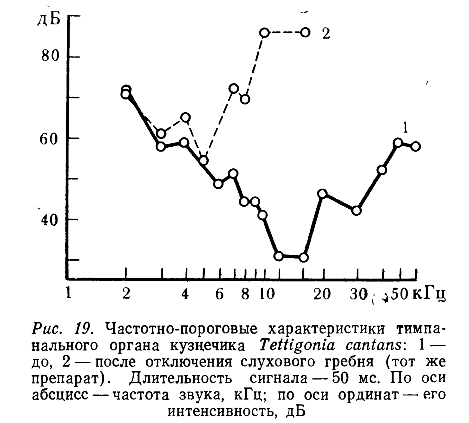
В следующей серии экспериментов перерезали нерв слухового гребня (см. рис. 13). Отключение этой группы сенсилл приводило к резкому изменению частотно-пороговых кривых (см. рис. 19). Оставшиеся сенсиллы промежуточного органа реагировали на звуки от 1 до 15 кГц, оптимум чувствительности наблюдался на 5 кГц, причем пороги реакции на этой частоте после операции не изменялись или повышались только на 1—5 дБ. Изменение частотно-пороговых характеристик тимпанального органа после отключения слухового гребня свидетельствует о том, что он состоит из двух отделов: малочувствительного низко частотного (пороговая интенсивность в зоне оптимума — около 55 дБ) и более чувствительного широкополосного (пороговая интенсивность в зоне оптимума — около 30 дБ).
Аналогичные результаты были получены при изучении двух Других видов кузнечиков. У D. verrucivorus промежуточный орган реагировал на звуки частотой от 2 до 15 кГц (оптимум — 5—6 кГц), а у РІг. pustulipes — от 2 до 10 кГц (оптимум — 7 кГц). Максимальная чувствительность неоперированных тимпанальных органов у этих видов наблюдается соответственно на 15 и 20—30 кГц.
Позднее, в результате исследования импульсной активности афферентных волокон у места их вхождения в I грудной ганглий У Decticus verrucivorus были зарегистрированы слуховые нейроны с разными частотными характеристиками (Rheinlaender, 1975). Их оптимальные частоты варьировали (у разных клеток) от 3 до 30 кГц. По некоторым функциональным характеристикам эти нейроны были разделены на две группы а и b. Ко второй из них отнесли рецепторы, обладающие меньшей чувствительностью к изменениям частоты, интенсивности и направления прихода звука.
Пытаясь связать эти данные с известными представлениями о строении тимпанальных органов, Райнлендер высказал предположение, что а-клетки соответствуют рецепторам слухового гребня, а b-элементы образуют промежуточный орган. Так как эта гипотеза находилась в очевидном противоречии с результатами наших опытов (Жантиев, 1971), мы провели эксперименты, впервые позволившие зарегистрировать у кузнечиков активность слуховых рецепторов непосредственно в тимпанальном органе (Жантиев, Корсуновская, 1978). Результаты этих опытов сводятся к следующему.
Слуховой гребень. В трех сериях экспериментов регистрировали активность сенсилл дистальной, средней и проксимальной частей слухового гребня. Нейроны, которые удалось зарегистрировать в дистальной части, отвечали на звук в диапазоне 9—60 кГц (рис. 20). Максимальная чувствительность наблюдалась в области 40—50 кГц. Минимальные пороги реакции достигали 60 дБ.
Вводя электрод в среднюю часть слухового гребня, теоретически можно было попасть в волокна, идущие от любых сенсилл, расположенных дистальнее места отведения. То есть наряду с элементами среднего отдела мы могли повторно зарегистрировать ответы упомянутых выше терминальных сенсилл. На практике этого, однако, не происходило. При введении электрода в среднюю часть слухового гребня регистрировались либо суммарные ответы многих нейронов, либо реакции одиночных клеток с иными частотными характеристиками. Одна из таких кривых с оптимумом на 14—16 кГц представлена на рис. 20. Объясняются эти результаты, по-видимому, тем, что в первом случае кончик электрода находился в тимпанальном нерве, а во втором — вблизи одного из аксонов до вхождения его в нерв. Следовательно, мы могли регистрировать ответы только тех сенсилл, которые находились на минимальном удалении от точки отведения.
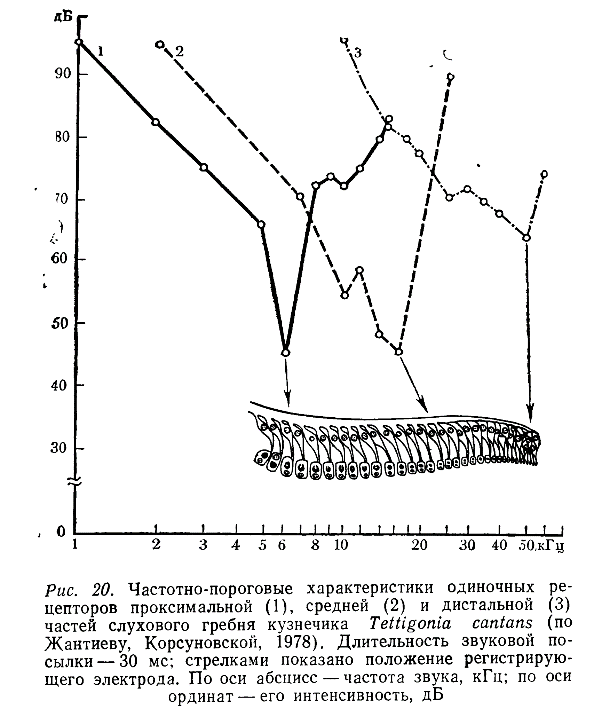
Аналогичные данные были получены при исследовании сенсилл проксимальной части слухового гребня. В этом случае регистрировались ответы нейронов, хорошо отличающихся по частотным характеристикам как от дистальных, так и от средних элементов. Диапазон воспринимаемых ими частот был ограничен 1 —17 кГц, а наибольшая чувствительность наблюдалась в области 6 кГц (рис. 20).
Эти результаты свидетельствуют о том, что сенсиллы слухового гребня различаются по частотным характеристикам. В проксимальной части расположены низкочастотные рецепторы, в дистальной — высокочастотные, а в средней части — сенсиллы, реагирующие на промежуточные частоты. Таким образом, мы получили прямое подтверждение высказывавшихся ранее предположений о существовании градиента в частотной избирательности рецепторов этого отдела тимпанального органа (Жантиев, 1971; Rheinlaender. 1975).
Ответы всех сенсилл слухового гребня были тоническими. Величина минимального латентного периода составляла 4 мс. При изучении зависимости числа импульсов в разряде от интенсивности стимула получены кривые, характерные для слуховых рецепторов других прямокрылых (рис. 21, А). Повышение интенсивности стимула над порогом приводило к возрастанию импульсации в диапазоне 20—25 дБ, затем наступало «насыщение».
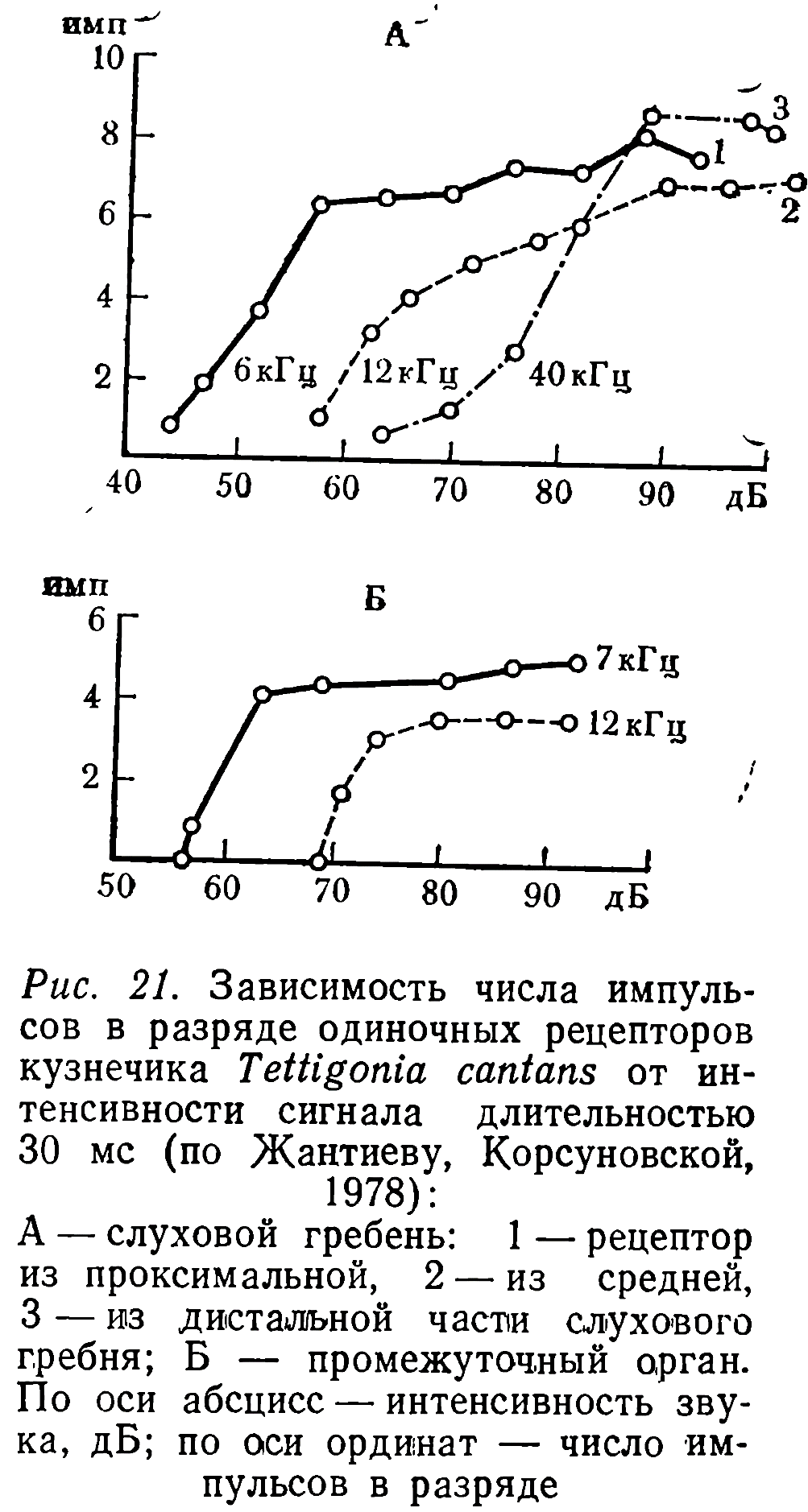
Нейроны промежуточного органа отвечали на звук в диапазоне от 2 до 12—14 кГц. Наибольшая чувствительность наблюдалась в области 5—7 кГц (рис. 22) (пороги — 60—80 дБ). Минимальный латентный период реакции составлял 6,7 мс. При повышении интенсивности стимула число импульсов в ответах изменялось так же, как и у нейронов слухового гребня, но динамический диапазон был ограничен 10—15 дБ (рис. 21, Б).
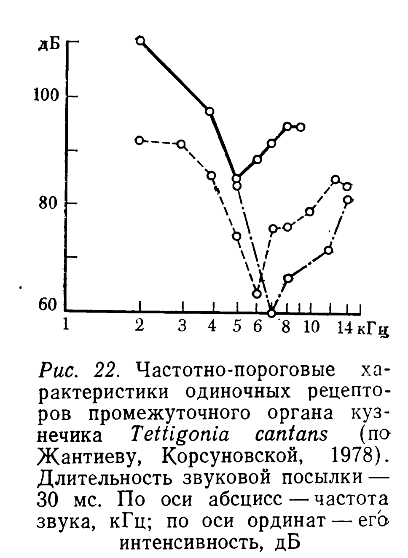
Эти данные свидетельствуют о том, что рецепторы промежуточного органа не обладают свойствами клеток типа b. В отличие от них сенсиллы промежуточного органа реагируют на звуки только в низкочастотной области спектра (оптимум — 5—7, а не 7—40 кГц), обладают V-образными частотно-пороговыми характеристиками и, кроме того, при повышении интенсивности стимула их импульсная активность быстро достигает предела (рис. 21). Нейронов типа b в тимпанальном органе Т. cantans мы не обнаружили.
Дополнительное исследование слуховых волокон в тимпанальном нерве Decticus verrucivorus показало, что деление рецепторов на группы а ч b лишено достаточных оснований (Kalmring et аЦ 1978).
При изучении функциональной организации тимпанальных органов кузнечиков много внимания уделяется не только рецепторному аппарату, но и вспомогательным структурам — в первую очередь тимпанальным мембранам и акустической трахее. На возможность участия последней в проведении звука указывалось еще в ранних морфологических работах (Gräber, 1876; Zeuner, 1936). Сравнительно недавно Люисом (Lewis, 1974 a;b) было установлено, что у кузнечика Homorocoryphus nitidulus (= Ruspolia differens) закрывание передних грудных дыхалец приводит к существенному повышению порогов реакции тимпанальных органов в широком диапазоне частот. Сопоставление этих данных с результатами анализа размеров акустической трахеи позволили предположить, что она представляет собой экспоненциальный рупор.
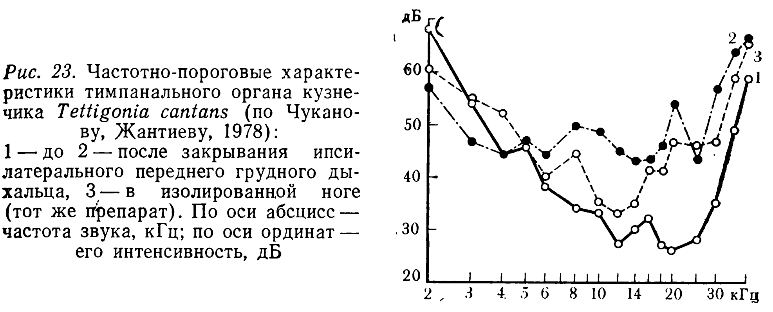
В процессе изучения слуховой системы Teiligonia cantans, издающей, в отличие от предыдущего вида, широкополосные сигналы, нами было установлено (Чуканов, Жантиев, 1978), что блокирование ипсилатерального дыхальца приводит к повышению порогов в диапазоне от 5 до 40 кГц (рис. 23, 1У 2). Опыты с укорочением акустической трахеи не приводили к смещению зоны оптимальной чувствительности в высокочастотную область (рис. 23, 3). Эти данные позволяют считать, что акустическая трахея Т. cantans обладает свойствами рупора, повышающего чувствительность рецепторов к звукам высоких частот.
Сходные результаты получены группой исследователей (Seymour et al., 1978; Michelsen, Larsen, 1978), изучавших акустические свойства тимпанальных органов у трех видов кузнечиков (Т. cantans, Т. viridissima и Ruspolia differens). Анализируя чувствительность тимпанальных органов и колебания мембран (с помощью метода лазерной виброметрии), они пришли к выводу что акустические трахеи выполняют роль рупоров, эффективности которых возрастает на высоких частотах. Ими было, в частности показано, что отношение внутреннего звукового давления к наружному при повышении частоты от 1 до 40 кГц возрастает в 3 раза.
Все эти данные находятся, однако, в противоречии с результатами исследований Нокке (Nocke, 1975), свидетельствующим! о том, что у австралийского кузнечика Acripeza reticulata аку стические трахеи обладают свойствами резонаторов, повышающих чувствительность тимпанальных органов только в области оптимальной частоты (8 кГц). Причина этих расхождений может быть обусловлена как спецификой объектов, так и различиям? в методике проведения опытов, поэтому для окончательного решения вопроса о функциональной роли акустических трахей у кузнечиков необходимы дополнительные сравнительные исследования.
Дата добавления: 2022-01-27; просмотров: 1053;