Ультраструктурная организация и функциональные характеристики слуховых рецепторов (хордотональных сенсилл)
Рецепторный аппарат тимпанальных и Джонстоновых органов насекомых состоит из хордотональных сенсилл, или сколопидиев. Несмотря на большое морфофункциональное разнообразие слуховых органов, их сенсиллы обладают сходными чертами организации, поэтому мы попытаемся дать их общую характеристику.
Ультраструктурная организация хордотональных сенсилл. Начало изучению субмикроскопического строения хордотональных сенсилл было положено в 1958—1960 гг. Грэем (Gray, Pumphrey, 1958; Gray, 1960), исследовавшим структуру интегументных сенсилл из тимпанального органа саранчи. Затем были изучены сенсиллы из тимпанальных органов кузнечиков (Жантиев, 1969; Жантиев, Перов, 1969), совок (Ghiradella, 1971), сверчков (Friedman, 1972; Young, Ball, 1974; Michel, 1974), цикад и клопов (Michel, 1975, 1977). Параллельно велись исследования этих рецепторов в Джонстоновых органах мух (Uga, Kuwabara, 1965), комаров (Risler, Schmidt, 1967; Belton, 1974), термитов (Howse, 1968), цикадок (Howse, Claridge, 1970) и жуков (Иванов, 1969).
Результаты этих работ показали, что в пределах однотипных органов сенсиллы имеют сходное строение. Примером рецепторов, характерных для тимпанального органа, могут служить сенсиллы из слухового гребня кузнечиков (рис. 17).
Каждая такая сенсилла представляет собой комплекс из 3—4 клеток. Основу его образует биполярный нейрон, от дендрита которого отходит довольно длинная ресничка, обладающая более или менее выраженным утолщением. В плазме реснички лежат 9 пар периферических фибрилл, связанных с корешковым аппаратом дендрита, а ее утолщенная часть обычно содержит три извитые нити. Снаружи дистальная часть дендрита и реснички окружена ребристым сколопсом — внутриклеточным продуктом обкладочной клетки. Дистальные концы ребрышек сколопса примыкают к конусовидной или колоколовидной шапочке — экстраклеточному образованию, пронизанному многочисленными полостями и канальцами. В центральной части шапочки расположен канал, в который входит дистальная часть реснички. Снаружи шапочку окружает шапочковая, или прикрепительная, клетка, обычно содержащая многочисленные тонофибриллы, а проксимальная часть дендрита заключена в складки фиброзной клетки и тяжи волокнистого материала, обеспечивающего прикрепление нейрона к близлежащим структурам.
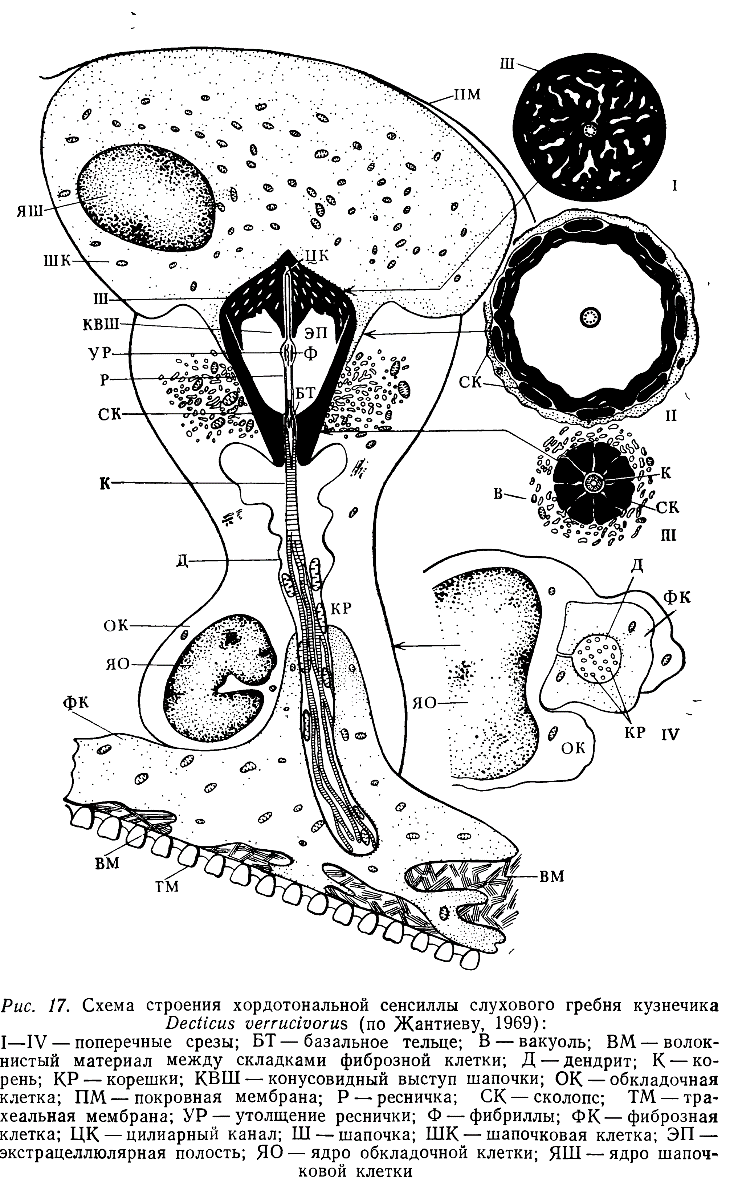
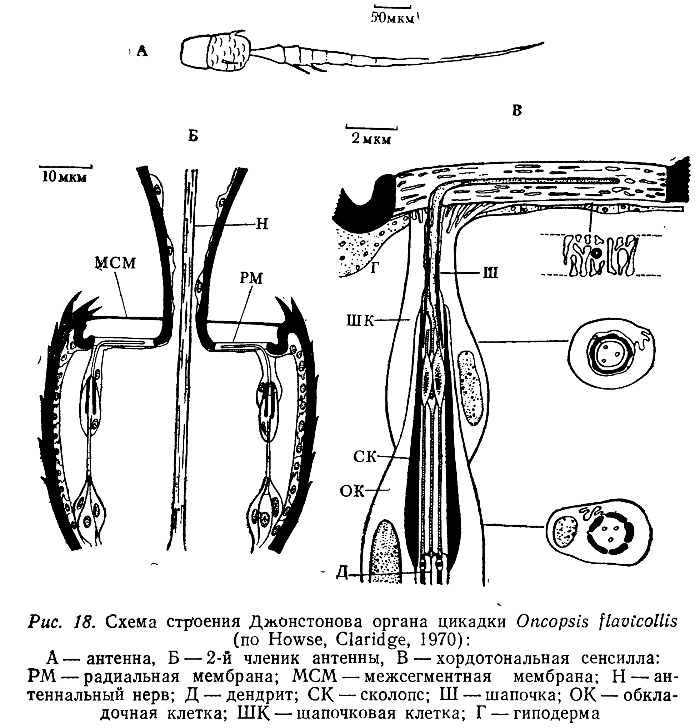
Джонстоновы органы состоят из модифицированных сенсилл, отличающихся от описанных главным образом наличием, двух (двукрылые) или трех (термиты, жуки, цикадки) нейронов и соответственно двух или трех ресничек в одном сколопсе. В последнем случае один из нейронов часто обладает в той или иной мере недоразвитой ресничкой, а у двух других ее дистальная часть бывает утолщена. Шапочка в этих сенсиллах превращена в длинный тонкостенный футляр, вершина которого обычно входит в сочленовную мембрану, соединяющую 2-й и 3-й членики антенны (рис. 18).
Функциональные характеристики хордотональных сенсилл. К настоящему времени получены функциональные характеристики отдельных рецепторов из тимпанальных органов кузнечиков (Rheinlaender, 1975; Жантиев, Корсуновская, 1978; Kalmring et ah, 1978), сверчков (Zaretsky, Eibl, 1978; Markovich, in: Eisner, Popov, 1978; Eibl, Huber, 1979; Чуканов и др., в печати), саранчовых (Michelsen, 1966, 1968, 1971а; Römer, 1976), совок (Suga, 1961; Roeder, 1966a; Adams, 1971), пядениц (Roeder, 1974), огневок (Agee, 1969c; Perez, Zhantiev, 1976) и водных полужесткокрылых (Weise, 1972; Arntz, 1975; Prager, 1973, 1976).
Механизм работы этих своеобразных рецепторов до сих пор остается неясным. Ранее высказывались предположения, что при стимуляции сенсилл происходит периодическое поперечное (Thurm, 1965) или продольное (Жантиев, 1969) сжатие реснички. В последнее время получены данные, свидетельствующие о том, что в ножных хордотональных органах стимуляция сенсиллы вызывает активное изгибание базальной части реснички (Moran et al., 1977).
При раздражении тимпанальных органов в нейронах хордотональных сенсилл возникает генераторный потенциал, амплитуда которого (1—3 мВ) при непрерывной стимуляции сохраняется на постоянном уровне по крайней мере в течение 1—2 с (Suga, 1960).
Уровень спонтанной импульсной активности рецепторов невысок и обычно колеблется от 1 до 15—20, реже — 40 имп./с.
Величина латентного периода реакции при прочих равных условиях зависит от места регистрации ответа. Непосредственно в тимпанальном органе у начала аксонов она составляет при околопороговых интенсивностях 8—16 мс, а при повышении уровня звука до 90—100 дБ сокращается до 2 мс. При регистрации ответов от волокон тимпанального нерва у места вхождения его в ганглий минимальные значения латентного периода приближаются к 5—6 мс.
По характеру ответов на звуковые посылки, сравнимые по длительности с естественными пульсами, хордотональные сенсиллы можно отнести к тоническим или фазно-тоническим рецепторам, так как у некоторых из них наблюдается небольшой оп-эффект. При длительной стимуляции частота импульсов сначала падает по экспоненте, а затем по прошествии 1—2 с обычно стабилизируется и удерживается на приблизительно постоянном уровне. Скорость адаптации может быть различной не только у разных видов насекомых, но и в пределах одного тимпанального органа.
Так, например, у совок клетки Ai и А2 хорошо различаются не только по чувствительности и амплитуде спайков, но и по скорости адаптации (Suga, 1961). В случае раздражения тимпанальных органов очень короткими звуковыми посылками или щелчками некоторые сенсиллы могут реагировать синхронно на стимулы, повторяющиеся с частотой 150—200 в секунду, но в большинстве случаев при частотах, превышающих 50 Гц, наступает альтернация, т. е. частота ответов снижается вдвое. В суммарных же ответах синхронизация прослеживается до 400 Гц (Autrum, 1960).
Чувствительность большинства слуховых рецепторов невелика: пороги реакции, регистрируемые обычными методами, лежат в области 20—40 дБ. Использование некоторых специальных методов позволяет обнаружить слабые ответы при уровне звука, не превышающем 7—10 дБ (Michelsen, 1971с). До сих пор считалось, что максимальной чувствительностью обладают сенсиллы Джонстонова органа (порог около 0 дБ) (Tischner, 1953), однако эти данные ;не были подтверждены в наших экспериментах.
Динамический диапазон, т. е. интервал интенсивностей, в пределах которого наблюдается изменение импульсации, составляет у большинства сенсилл из тимпанальных органов 30—40 дБ. Если для раздражения используются звуковые посылки, превышающие по длительности 5 мс, то частота импульсации в определенных пределах возрастает пропорционально логарифму звукового давления, а затем наступает «насыщение» или стабилизация импульсации.
Однако при изучении тимпанальных органов огневок мы наблюдаем последующее снижение уровня импульсной активности (Perez, Zhantiev, 1976). Аналогичное явление отмечалось у некоторых совок (Suga, 1961) и видов из семейства Ctenuchidae (Coro et al., 1977). Если же рецепторы стимулируют очень короткими посылками, то частота импульсов, являясь функцией длительности стимула и его интенсивности, возрастает непрерывно, и насыщения не наступает (Adam, 1971). Максимальный уровень импульсации обычно не превышает 300—400 имп/с, но в некоторых случаях (кузнечики, совки) достигает 600 или 700 имп/с.
При изучении функций слуховых органов насекомых большое внимание всегда уделялось их частотным характеристикам. На первых этапах этих исследований была выдвинута гипотеза, отрицавшая способность слуховых рецепторов дискриминировать звуки по частоте. Иначе говоря, считалось, что, в отличие от позвоночных, у насекомых все сенсиллы слухового органа «настроены» на один диапазон частот (Pumphrey, Rawdon-Smith, 1939). В ходе дальнейших работ, проводившихся на представителях разных отрядов насекомых, были получены многочисленные подтверждения этой гипотезы. Однако в последние годы удалось установить, что тимпанальные органы прямокрылых имеют более сложную функциональную организацию, чем предполагалось ранее, и, в частности, содержат группы сенсилл, по-разному реагирующих на звуки разных частот.
Подробнее этот вопрос будет расмотрен в следующем разделе. Здесь же мы только отметим, что ширина диапазона частот, на которые может реагировать отдельная сенсилла, колеблется у разных видов от 10—12 до 50 кГц, а острота частотно-пороговой характеристики (Q), определяющаяся отношением оптимальной частоты (f0) к ширине полосы ![]() при уровне интенсивности, на 20 дБ превышающем порог, варьирует от 0,6 до 2,4.
при уровне интенсивности, на 20 дБ превышающем порог, варьирует от 0,6 до 2,4.
Дата добавления: 2022-01-27; просмотров: 894;