Акустические системы связи насекомых
Согласно определениям, принятым в кибернетике, любая система связи включает источник информации, передатчик, канал связи, приемник (дешифратор) и адресат. Все эти элементы, несомненно, имеются в акустических системах связи насекомых (рис. 1).
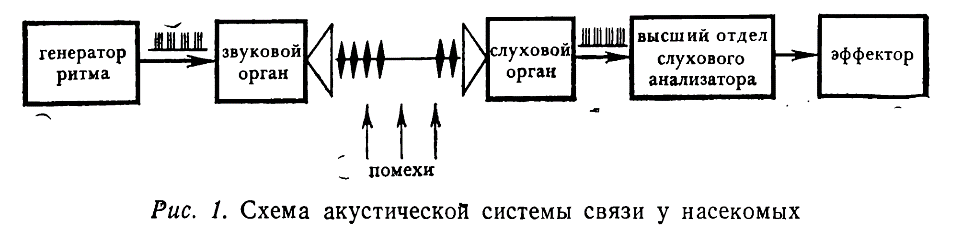
Источниками информации в этом случае служат находящиеся в ЦНС осцилляторы, генерирующие видоспецифические залпы импульсов; поступающая отсюда информация перекодируется в звуковые посылки с помощью звуковых органов (передатчик) и распространяется далее в воздушной, водной или твердой среде (канал связи).
Функции приемников у насекомых выполняют разнообразные слуховые органы, преобразующие звуковые колебания в нервные импульсы, а адресатом являются те отделы ЦНС, которые используют поступающую акустическую информацию для управления определенными поведенческими актами. Важной особенностью акустических коммуникационных систем насекомых следует считать высокий уровень помех в канале связи, заставляющей предполагать наличие в их ЦНС специальных механизмов, обеспечивающих выделение полезного сигнала из шумов, поступающих на вход приемника.
Механизмы звукоизлучения. Насекомые издают звуки с помощью пяти основных механизмов:
1. трением подвижно сочлененных склеритов (фрикционный или стридуляционный);
2. вибрацией специализированных мембран (тимбальный);
3. пропусканием воздуха через отверстия (пневматический);
4. ударением различными частями тела по субстрату (ударные механизмы);
5. вибрацией крыльев и связанных с ними структур.
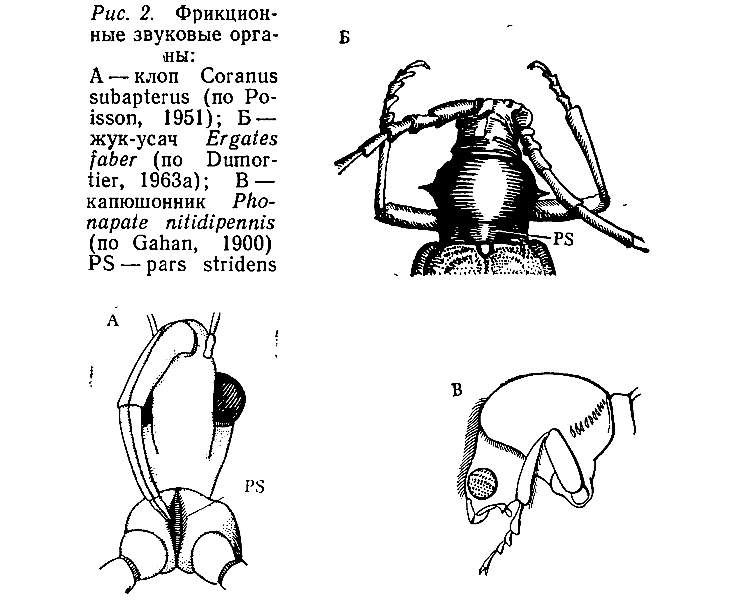
Фрикционные механизмы. Звуковые органы фрикционного типа обычно состоят из двух основных частей — острого канта (plectrum) и зазубренной пластинки (pars stridens), расположенных на соприкасающихся, подвижно сочлененных склеритах. В процессе эволюции эти органы возникают у представителей разных отрядов и даже семейств независимо и неоднократно, поэтому нет ничего удивительного в том, что они занимают разное положение на теле насекомого. По этому признаку могут быть выделены органы приблизительно 25 типов (Dumortier, 1963а).
Примером сравнительно простых фрикционных аппаратов могут служить стридуляционные органы некоторых жуков и клопов. У жуков-усачей (Cerambycidae) на медиальной части среднеспинки имеется покрытая тонкими ребрышками площадка, по которой скользит задний край переднеспинки (рис. 2, Б). Клопы-хищнецы (Reduviidae) издают звуки с помощью хоботка, трущегося о зазубренный участок переднегруди (рис. 2, А), а некоторые капюшонники (Bostrychidae) используют для этой цели бугорки на переднеспинке и небольшие напильнички на вершинах передних бедер (рис. 2, В).
Максимальной сложности звуковые органы фрикционного типа достигают у прямокрылых. У большинства саранчовых (Acridoidea) на внутренней стороне задних бедер находится продольный ряд бугорков, который соприкасается с одной из жилок надкрылий. Реже наблюдается обратная картина: бугорки располагаются на надкрыльях, а острые канты — на бедрах. Во время стри- дуляции насекомые поочередно поднимают и опускают задние ноги, в результате чего возникают вибрации надкрылий, излучающих сравнительно громкие звуки с довольно широким спектром. У кузнечиковых (Tettigonioidea) на внутренней поверхности левого надкрылья расположена зазубренная жилка, которая во время стридуляции трется о внутренний край правого надкрылья (рис. 3).
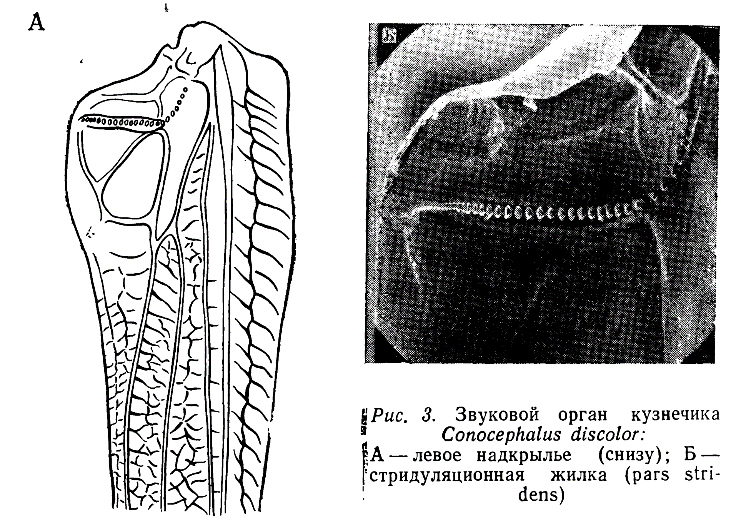
Аналогичная жилка имеется и на правом надкрылье, но она, как правило, не функционирует. К ней примыкает тонкая мембрана, так называемое «зеркальце», представляющее собой более или менее увеличенную кубитальную ячейку. В случае частичной редукции надкрылий звуковой аппарат обычно полностью сохраняется. У сверчковых (Crylloidea), в отличие от кузнечиковых, функционирует pars stridens на правом надкрылье, а зеркальце расположено в дистальной части надкрылий. У медведок (Gryllotalpidae) зеркальце отсутствует.
У подавляющего большинства прямокрылых звуковыми органами обладают только самцы. У самок они либо отсутствуют, либо бывают развиты в значительно меньшей степени, чем у самцов.
При изучении функций фрикционных органов основное внимание уделяется механическим свойствам звукоизлучающих структур, особенностям движения частей тела, на которых они расположены, и работе нервно-мышечного аппарата, приводящего их в движение.
В результате тщательного исследования звуковых органов кузнечиков установлено, что основным источником колебаний служит жесткая рамка, образованная жилками, окружающими «зеркальце». Если частота ударов plectrum по зубчикам pars stridens соответствует собственной частоте колебаний рамки и прилегающих к ней снаружи участков надкрылий, то эта «резонансная система» излучает более или менее чистые тональные сигналы (кузнечики рода Homorocoryphus). Если же такое соответствие не наблюдается (большинство кузнечиков Tettigonüdae), то каждый удар зубчика вызывает свободные колебания системы. Обычно они частично или полностью затухают к началу следующего удара (рис. 4, А), поэтому звук распадается на более или менее отчетливые щелчки («нерезонансная система») (Morris, Pipher, 1967; Bailey, 1967; 1970; Bailey, Broughton, 1970). К последнему типу относятся и звукоизлучающие системы саранчовых (Eisner, 1974).
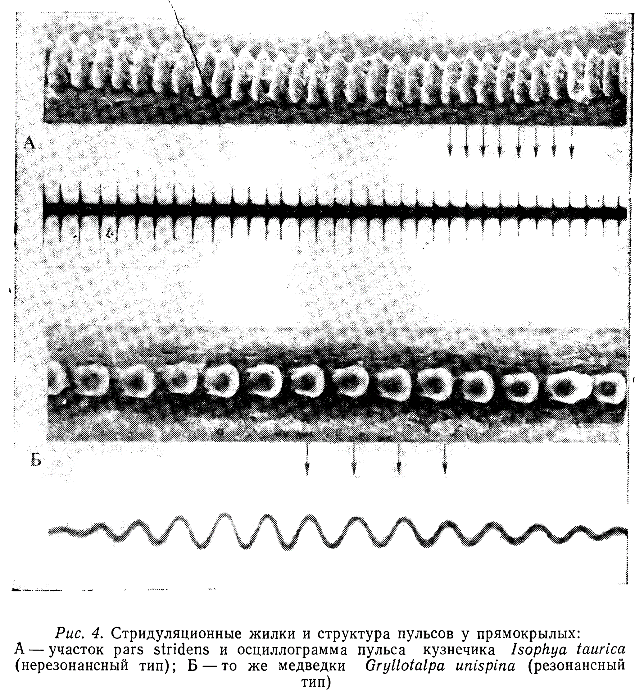
У сверчков главной излучающей структурой является так называемая «лировидная часть» надкрылий; ее резонансная частота совпадает с основной частотой призывного сигнала (около 5 кГц) (Nocke, 1971). Приблизительно так же функционируют и звуковые органы медведок (Bennet-Clark, 1970) (рис. 4, Б). У большинства длинноусых прямокрылых (Ensifera) звуковые посылки возникают при сдвигании надкрылий, но во многих случаях их раздвигание также сопровождается эмиссией звука меньшей амплитуды (рис. 7).
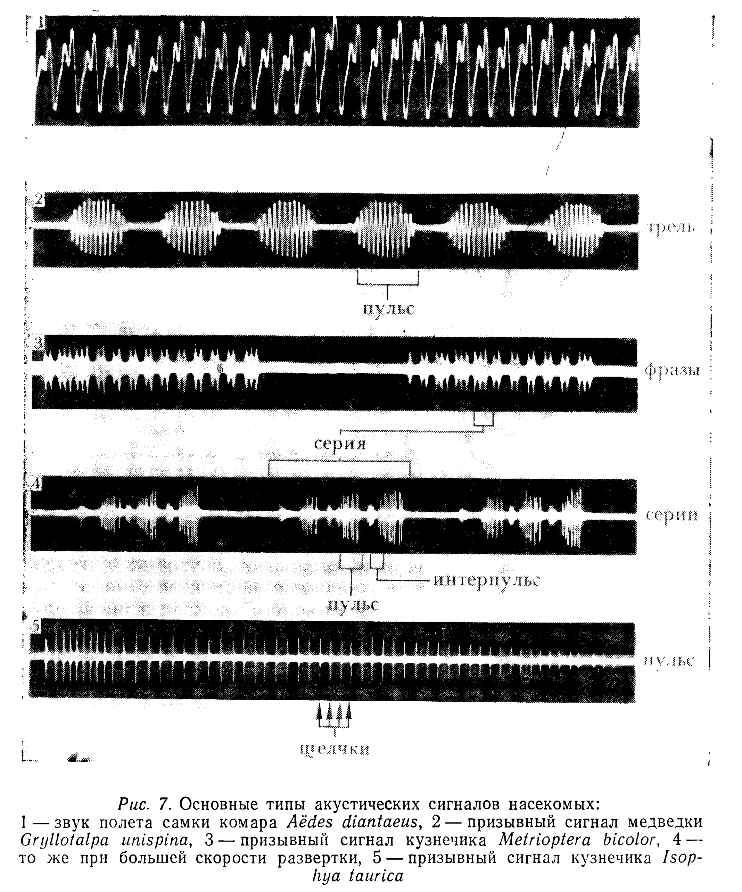
Использование специальных методов регистрации движений надкрылий у кузнечиков и задних ног у саранчовых позволило обнаружить интересные модификации в работе их звуковых органов. Так, например, у некоторых видов кузнечиков смыкание надкрылий происходит в два этапа с закономерно изменяющейся скоростью, причем не каждое смыкание сопровождается эмиссией звука (Walker, Dew, 1972; Walker, 1975а).
У многих саранчовых усложнение ритмической организации сигналов достигается за счет комбинации стридуляционных движений разных типов: поднимания, опускания и быстрых вибраций бедер. При этом парные звуковые органы могут работать с некотором фазовым сдвигом или даже по разным программам (Eisner, 19/3, 1974; Eisner, Popov, 1978).
Нервно-Мышечный аппарат, приводящий в движение фрикционные звуковые органы, хорошо изучен у сверчков, саранчовых и кузнечиков (обзоры: Eisner, Huber, 1973; Huber, 1974b, 1975; Eisner, Popov, 1978).
Анализ электромиограмм, зарегистрированных во время стридуляции, свидетельствует о том, что нервная система обеспечивает не только четкую координацию работы мышц, приводящих в движение надкрылье или ногу, но и последовательность срабатывания моторных единиц в пределах одной мышцы, а также фазовые соотношения в движениях парных органов. Так, например, у сверчков при издавании призывного сигнала поочередно сокращаются две группы мышц-антагонистов, сводящих и разводящих надкрылья (рис. 5, А). Причем мышцы в пределах каждой группы (синергисты) срабатывают почти синхронно (Kutsch, 1969). Приблизительно такая же картина наблюдается у быстро стридулирующих саранчовых, но у видов, сравнительно медленно поднимающих и опускающих бедра, соответствующие мышцы сокращаются в определенной последовательности (рис. 5, Б) (Eisner, 1968, 1975).
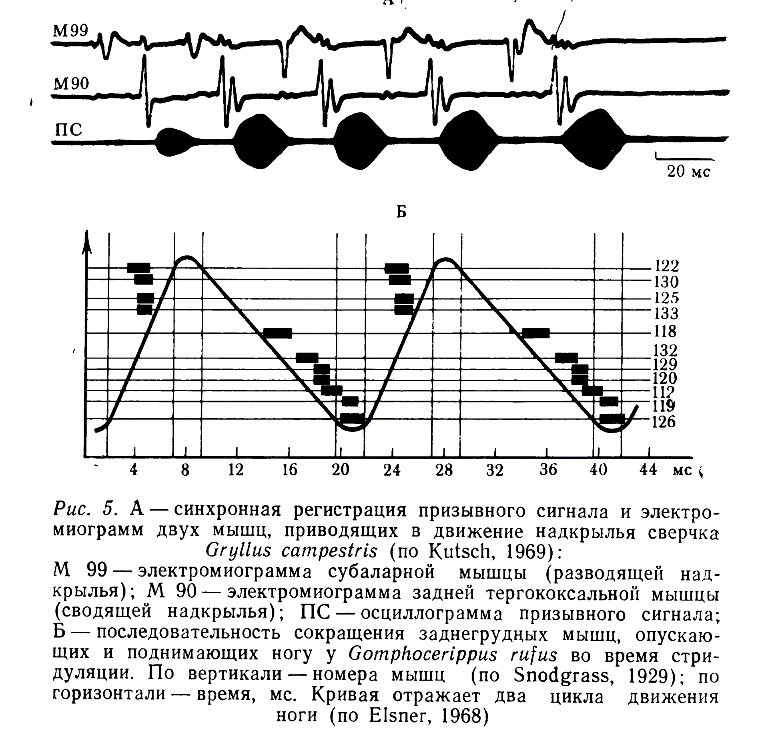
Разнообразные проприоцепторы, по-видимому, не оказывают непосредственного влияния на ритм стридуляцни. Сразу после их отключения в сигналах или импульсации мотонейронов не удается обнаружить заметных изменений (Eisner, Huber, 1969; Kutsch, Huber, 1970; Möss, 1971). Однако через несколько дней после такой операции во временной организации сигнала все же проявляются некоторые искажения (Lindberg, Eisner, 1977), поэтому есть основания думать, что функционирование этих рецепторов необходимо для нормальной работы звуковых органов.
Различные визуальные, ольфакторные или механические стимулы могут оказывать значительное влияние на акустическое поведение насекомых. Координация их звуковой активности с общим поведением в конкретной ситуации осуществляется интегративными центрами надглоточного ганглия. Раздражая электротоком определенные области этого ганглия у закрепленных или свободно двигающихся сверчков, можно получить один из трех внутривидовых сигналов: призывный, агрессивный или прекопуляционный (см. рис. 9).
Изменение частоты повторения или амплитуды стимула приводит к замене одного сигнала другим, но не вызывает искажений основных временных параметров каждого из них (Huber, 1960, 1962, 1963; Otto, 1969, 1971). Таким образом, функции надглоточного ганглия сводятся в данном случае к запуску и регуляции продолжительности звучания каждого из трех сигналов, а их стабильные видоспецифические параметры целиком определяются звуковым центром в грудных ганглиях.
Функциональная организация этого центра изучена недостаточно, но результаты внутриклеточной регистрации активности мезоторакальных мотонейронов сверчка дают основание предполагать, что их активность контролируется двумя генераторами ритма — быстрым и медленным. Первый задает ритм элементарных звуковых посылок (пульсов), а второй — определяет длительность и частоту повторения их группировок (серий) (Bentley, 1969 а, b).
Тимбальные и Пневматические механизмы. Тимбальные звуковые органы имеются у многих представителей Homoptera, некоторых Heteroptera и Lepidoptera. Долгое время считалось, что в отряде Homoptera этими органами обладают только самцы крупных цикад семейства Cicadidae, но впоследствии они были обнаружены и у мелких форм, относящихся ко многим другим семействам (Eurimelidae, Cicadellidae, Cercopidaе и др.). Причем оказалось, что у этих насекомых зву&и издают не только самцы, но и самки. У цикад тимбальные органы располагаются в I брюшном сегменте. Основу этого органа образует выпуклая мембрана, к которой изнутри прикрепляется мощная тимбальная мышца и небольшая группа вспомогательных тензорных волокон (рис. 6).
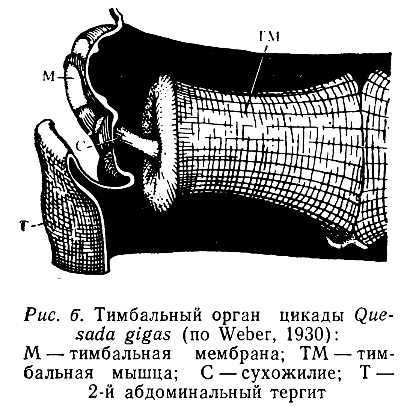
Механизм работы такого аппарата сравнительно прост. При сокращении тимбальной мышцы упругая мембрана прогибается внутрь, а при ее расслаблении возвращается в исходное положение. В результате этого генерируется 2 пульса, которые усиливаются различными резонирующими структурами й в первую очередь большими воздушными полостями (трахейными мешками), занимающими значительную часть брюшка (Pringle, 1954; Hagiwara, 1956).
Тензорные мышцы, по-видимому, регулируют натяжение (и кривизну) мембраны и тем самым оказывают определенное влияние на интенсивность и частоту повторения пульсов (Pringle, 1954; Aidley, 1969; Simmons, Young, 1978). Цикады, так же как и прямокрылые, могут издавать тональные и шумовые звуки. Основные или доминирующие частоты их сигналов обычно соответствуют резонансной частоте тимбальной мембраны. На ширину спектра излучаемых частот определенное влияние оказывают трахейные мешки, выполняющие (по крайней мере у некоторых видов) роль резонаторов.
Для этих насекомых характерна очень высокая частота повторения пульсов. Повышение частоты их повторения достигается у разных видов с помощью двух механизмов: а) тимбальные мышцы отвечают на один импульс мотонейрона несколькими сокращениями (миогенный ритм) (Pringle, 1954; Hagiwara et al., 1954); б) обе мышцы отвечают на каждый импульс одним сокращением (нейрогенный ритм), но работают поочередно, в результате чего ритм удваивается (до 800 пульсов в секунду). У многих видов цикад тимбальная мембрана имеет ребристую структуру, поэтому при однократном сокращении мышцы, она прогибается в несколько этапов, генерируя серию (до 6) звуковых импульсов, образующих один сложный іп-пульс (Moore, Sawyer, 1966; Reid, 1971; Young, 1972a; Попов, 1975). Каждая тимбальнай мышца иннервируется одним мотонейроном. Видоспецифический ритм импульсации этих клеток определяется осцилляторами (пейсмекерами) звукового центра (Hagiwara, Watanabe, 1956; Свидерский, 1969, 1973; Simmons, 1977).
У клопов-шитников (Pentatomidae) небольшие тимбальные органы располагаются в двух первых сегментах брюшка. Тим- балы чешуекрылых из семейств Arctiidae и Ctenuchidae представляют собой несколько модифицированные метэпистерны. При сокращении базалярных мышц поверхность этих склеритов волнообразно изгибаемся, в результате чего излучается серия коротких звуковых импульсов. Их число обычно соответствует количеству поперечных ребрышек на поверхности тимбальной мембраны. Расслабление мышц и выпрямление мембраны сопровождается эмиссией второй серии импульсов. У видов с гладкими тимбалами при однократном сокращении мышцы генерируются только два пульса (Biest et al., 1963; Fenton, Roeder, 1974; Fullard, Fenton, 1977).
Пневматические механизмы. Неоднократно высказывались предположения, что насекомые могут издавать звуки, пропуская воздух через дыхальца. Некоторые исследователи считают, что именно таким способом издают звуки жуки-плавунцы и их личинки, пчелиные матки и двукрылые. Более определенно известно, что такой механизм функционирует у тараканов рода Gromphadorhina. Эти насекомые издают шипящие звуки, выпуская воздух из пары специализированных брюшных дыхалец (Dumortier, 1965; Nelson, 1979).
Бражник «мертвая голова» (Acherontia atropos) может всасывать воздух через хоботок в глотку, в результате чего возникают вибрации эпифаринкса, дающие колебания сравнительно низких частот (5—15 кГц). При выпускании воздуха эпифаринкс приподнимается и создаются условия для генерации высокочастотных звуков (3,5—30 кГц) (Prell, 1920; Busnel, Dumortier, 1959).
У некоторых саранчовых выделение пены из грудных дыхалец сопровождается хорошо слышимым звуком. Аналогичное явление наблюдается и у чешуекрылых (Arctiidae, Zygaenidae) с той лишь разницей, что у них пенистые вещества выделяются из специальных желез. Механизм звукоизлучения в этих случаях изучен недостаточно. У жуков-бомбардиров (Brachinus, Carabidae) звук возникает в результате «взрыва», образующегося при выбрызгивании из анального отверстия легко испаряющейся жидкости, содержащей азотнокислые соли и окись азота.
Ударные механизмы. Многие насекомые издают звуки, ударяя по субстрату различными частями тела. Особенно успешно этот способ звуковой сигнализации используется насекомыми, живущими в древесине. Так, некоторые жуки-точильщики (Anobiidae), ударяя головой о стенки ходов, издают звуки, напоминающие тиканье часов (так называемые «часы смерти»).
Хорошо известно, что солдаты термитов (Isoptera) оповещают членов семьи об опасности, ударяя (часто синхронно) по стенкам гнезда головой. Приблизительно такой же способ сигнализации наблюдается и у некоторых муравьев (Camponotus, Polyrhachis). Личинки шершней (Vespa) царапают ротовым аппаратом стенки ячеек. Многие сеноеды (Psocoptera), веснянки (Plecoptera), вислокрылки (Megaloptera), скорпионовые мухи (Mecoptera), чешуйчатые сверчки (Mogoplistidae), жуки (Tenebrionidae) издают звуки, постукивая по субстрату кончиком брюшка. При этом некоторые из них для усиления звука специально прикасаются вибрирующим брюшком к сухим листьям и другим резонирующим предметам. Реже для такой сигнализации используются конечности. Известно, например, что некоторые саранчовые (Oedipodinae) стучат по земле задними лапками.
Механизмы, связанные с работой крылового аппарата.Работа крылового аппарата насекомых почти всегда сопровождается борее или менее отчетливым звуком. Его источником могут служить как сами крылья, так и стенки тела, вибрирующие под воздействием крыловых мышц. Некоторые насекомые издают звуки не только во время полета, но и при других видах активности. Особенно интенсивно этот механизм звукоизлучения используется в гнездах общественных перепончатокрылых — пчел, шмелей и ОС.
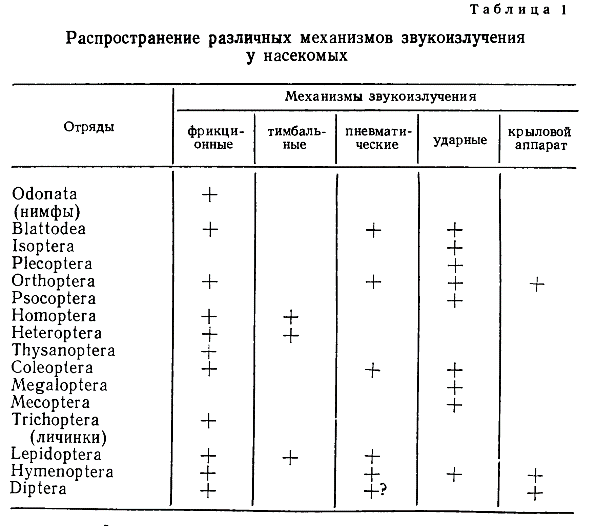
При анализе звуков, возникающих в полете, иногда приходится учитывать возможность работы дополнительных механизмов — фрикционных (Oedipodinae) или тимбальных (Arctiidae, Ctenu- chidae). Некоторое представление о распространении разных механизмов звукоизлучения у насекомых дает табл. 1.
Дата добавления: 2022-01-27; просмотров: 1261;