Полярные океаны с точки зрения биологии и экосистем
В предыдущем разделе перечислен ряд причин, по которым процессы в Северном Ледовитом и Южном океанах отличаются от процессов в других океанах и по которым они так несхожи между собой. И те и другие отличия — важнейшие условия для возникновения форм жизни в полярных океанах, а происходящие там физические процессы создают основу для изучения их биологии и экосистем. Прежде чем приступить к изучению данных вопросов, сделаем обзор этих различий и процессов.

Океанология через дырку во льду! Большая часть того, что мы знаем о Северном Ледовитом океане, получена в результате опускания приборов в отверстия, проделанные в ледяных полях.
1. Полярная океанология возникла относительно недавно и менее развита, чем наука об океанах умеренных и тропических широт. В значительной мере это обусловлено удаленным географическим положением и большой стоимостью экспедиций в высокие широты. Наша база данных для обоих полярных океанов скудна. Сведения о Северном Ледовитом океане редки из-за того, что постоянный ледяной покров в центральной части бассейна не пропускает никакие исследовательские суда, кроме ледоколов. Об антарктических морях известно очень мало из-за их удаленности и высокой стоимости перевозок и снабжения. Кроме того, стараться получить какие-либо данные зимой — значит подвергать судно опасности вмерзнуть в паковый лед и быть раздавленным льдинами, когда зимние штормы взламывают ледяные поля.
2. Поскольку влияние температуры на плотность холодной воды ничтожно по сравнению с влиянием солености, процессы, контролирующие приток пресной воды в полярные океаны, определяют также их циркуляцию, особенно в Северном Ледовитом океане.
3. Особый характер удаления ионов растворенных солей из кристаллической структуры воды в твердом состоянии — физическая основа для огромного влияния сезонного замерзания и таяния морского льда на циркуляцию воды в океанах, особенно в Южном океане.
4. Северный Ледовитый и Южный океаны сообщаются с другими океанами совершенно по-разному: Северный Ледовитый океан изолирован, соединяется на большой глубине только с Атлантикой и лишь через один пролив между Шпицбергеном и Гренландией; воды Южного океана с его Циркумполярным течением смешиваются с водами всех крупных океанических бассейнов и на всех уровнях — от поверхности до глубоководных абиссальных впадин.
5. Важное значение имеет разная стратификация вод Северного Ледовитого и Южного океанов по вертикали. Воды Северного Ледовитого океана резко расслоены, при этом в поверхностном слое воды поступление питательных веществ из более глубоких слоев ограничено, и поэтому первичная продукция в нем мала. Южный океан не имеет четкой стратификации, в нем происходит апвеллинг больших масс воды, питательные вещества в эвфотической зоне практически не ограничены и первичная продукция велика.
И следствием этих различий являются две главные стороны жизни в полярных океанах: почти полное отсутствие видообразования в Северном Ледовитом океане и громадная биологическая продуктивность в Южном океане.
Слабое видообразование в Северном Ледовитом океане. Одна из самых странных и удивительных особенностей жизни в полярных океанах — несомненно, малое число видов как растений, так и животных по сравнению с гораздо большим разнообразием видов в морях умеренных и тропических широт. Нигде это не носит столь поразительного характера, как в Северном Ледовитом океане. Отрывок из работы Данбара [4] очень хорошо характеризует ситуацию в Северном Ледовитом океане.
«Бедность фауны и флоры Арктики хорошо известна. Напри-мер, здесь обитает, вероятно, всего 25—30 видов рыб, которые принадлежат именно водам Северного Ледовитого океана (из тысяч видов, относящихся ко всем океанам), из морских планктонных копепод (1500 видов во всех океанах) в Северном Ледовитом океане встречается лишь около 40 видов, из хетогнат (всего в Мировом океане известно свыше 30 видов) в Арктике обнаружено только два вида, а примерно из 85 известных видов эвфаузиид в субарктических водах (например, в Баренцевом море) найдено четыре и ни одного — в арктических (полярных) водах.
Для четырех групп шельфового бентоса (ракообразные, иглокожие, моллюски и полихеты) характерно уменьшение числа видов с приближением к арктическим условиям. Отсюда следует, что малое разнообразие видов характерно для всей фауны Северного Ледовитого океана».
Пока мы не изучим экосистему Северного Ледовитого океана достаточно хорошо, чтобы объяснить вопрос видообразования, мы не поймем как следует никакой экосистемы. Был предложен целый ряд факторов в качестве «причин того, почему» в Северном Ледовитом океане так мало видов.
Низкий уровень первичной продукции. Низкие значения первичной продукции, полученные в центральной части Северного Ледовитого океана, резко отличаются от более высокой продуктивности некоторых субарктических морей (таких, как Берингово море) и Южного океана. Например, Данбар [4] указывает для первичной продукции центральных вод Северного Ледовитого океана величину 0,6 г С/(м2-год); сравните ее с такими значениями, как 0,63 г С/(м2-сут) для центральной части Берингова моря (7], 1 г С/(мг-сут) в море Уэдделла в Антарктике или с величиной 300 г С/(м2-год), приведенной в табл. 1.2 для областей апвеллинга в целом.
Обратите внимание на единицы времени в этих данных о продукции — год и сутки. Дело в том, что ученые просто не в состоянии проникнуть в Берингово море или в море Уэдделла в зимнее время, поэтому мы не можем вычислить для них годовое среднее значение. В противоположность этому в центральной части Северного Ледовитого океана на льдинах были организованы дрейфующие станции с научным персоналом, работающим круглый год, научно-исследовательские рейсы в районы апвеллинга также проводятся круглогодично.
Одна из причин такой низкой продуктивности вод Северного Ледовитого океана состоит, вероятно, в сильном плотностном расслоении воды, препятствующем перемешиванию с более глубокими водами, что снижает пополнение поверхностной эвфотической зоны питательными веществами. И, как говорилось раньше, такое резкое расслоение воды в значительной мере обусловлено обильным речным стоком в этот бассейн, ограниченным перемешиванием с водами соседних океанов и процессом образования и таяния морского льда, который заставляет соли перемещаться в толще воды вниз.
Другая причина — задерживание света самим ледяным покровом. На фотографии пакового льда (рис. 20.8) видно, что белый, покрытый снегом лед сильно отражает солнечное излучение.

Открытые воды, где поглощается большая часть солнечных лучей, имеют почти черный цвет, но на такие щели в перемещаемом ветром подвижном паковом льду приходится лишь 10% площади поверхности многолетнего ледяного покрова. Интенсивность света в воде непосредственно под плавучим льдом составляет лишь небольшую часть интенсивности света, падающего на поверхность льда.
Под лед, покрытый снежным чехлом, проникает, вероятно, менее 1% света, достигающего поверхности, и после таяния снега это значение возрастает до нескольких процентов. Таким образом, ледяной покров резко ограничивает необходимое для фотосинтеза освещение в находящихся под ним водах.
Еще одно условие, влияющее на величину первичной продукции в покрытых льдом водах, — время массового размножения («цветения») фитопланктона. В средних широтах оно начинается в конце марта, в то время года, когда быстро увеличивается продолжительность дня. Однако в высоких широтах солнце не поднимается высоко над горизонтом. Вместе с ледяным покровом это задерживает начало «цветения». На рис. 20.9 показано, как оба' этих условия действуют вместе в прибрежных зонах моря Бофорта; взрывной рост фитопланктона начинается только в июле, после того как ледяной покров в значительной степени растает.
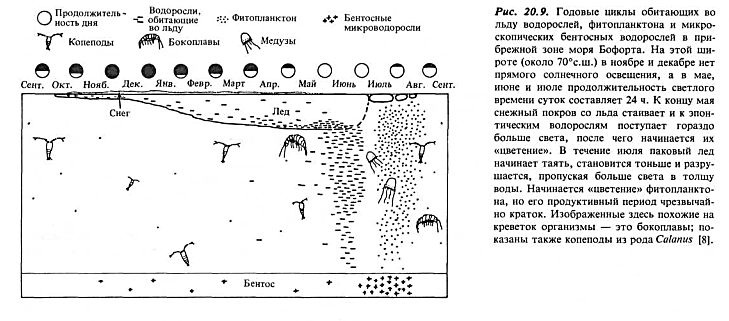
Еще одна особенность роста растений в покрытых льдом морях (рис. 20.9) — удивительная способность водорослей развиваться внутри самого льда. Эти водоросли назвали эпонтическими, что означает «растущие вне воды», хотя, строго говоря, они все же живут в воде, несмотря на то что находятся внутри льда. «Цветение» этих водорослей начинается в апреле и достигает максимума, когда стаивает снежный покров, приблизительно 1 июня. По мере таяния льда водоросли попадают в воду. Сообщества эпонтических водорослей представлены главным образом пеннатными диатомеями и микрофлагеллятами, последние имеют размеры примерно 6 мкм.
Эпонтические водоросли могут составлять до 25% первичной продукции. Но как они растут во льду и как могут способствовать питанию растительноядных и животных более высоких трофических уровней, если они закованы в лед? Ответы на эти вопросы все еще не найдены, но известно, что нижняя сторона пакового льда — это не твердая поверхность, а скорее похожее на губку скопление пластинок льда, перемешанных с жидкостью. Эпонтические водоросли обитают внутри такого слоя на поверхностях раздела льда и жидкости. Здесь они осуществляют газообмен и получают питательные вещества из жидкой воды, точно так же как любой фитопланктон. Некоторые водоросли оказываются в ловушках (карманах) вместе с рассолами во время быстрого замерзания воды и могут жить таким образом долгое время.
Эпонтическими водорослями питаются главным образом бокоплавы (амфиподы). По форме они напоминают эвфаузиид (рис. 20.9), а размеры их достигают нескольких сантиметров. Как-то я сидел у края отверстия, вырубленного во льду, с опущенной в воду под лед электрической лампочкой, которая освещала его нижнюю поверхность, и наблюдал, как вдоль этой поверхности носятся бокоплавы. В водах Северного Ледовитого океана видов этих животных больше, чем планктонных копепод, что наводит на мысль о важной роли обитающих во льду водорослей для арктической экосистемы. Более высокоорганизованные животные — рыбы, которые питаются амфиподами, а также птицы — представляют собой часть этой экосистемы. Нижняя поверхность льда становится своего рода «перевернутым» субстратом, относительно твердым, на котором развивается особое сообщество организмов.
Очень короткий сезон продуктивности. Из рис. 20.9 мы делаем вывод, что фаза высокой продукции в годовом цикле Северного Ледовитого океана весьма коротка по сравнению с циклом, изображенным на рис. 12.4 для океанов умеренных широт. Это должно оказывать сильное влияние на жизненные циклы растительноядных; им может не хватать времени, чтобы вырасти до половозрелого состояния за один сезон, и совпадение во времени личиночных стадий с достаточным количеством пищи может представлять проблему. Например, продолжительность жизни крупных копепод в Северном Ледовитом океане составляет по крайней мере один год, и, возможно, такая особенность развилась именно в этой экосистеме для того, чтобы воспроизводство потомства в начале второго года совпадало с новым периодом максимума продукции фитопланктона.
Продолжительный жизненный цикл у растительноядных видов приводит к тому, что внутри экосистемы запасается больше энергии, следовательно, существующая в каждый момент наличная биомасса растительноядных сравнительно велика. Отчасти по этой причине субарктические моря, такие как Берингово, необычайно богаты рыбой. В субарктических широтах паковый лед летом тает быстрее, а период образования первичной продукции продолжается несколько дольше, чем в Центральной Арктике, но тем не менее ее изменения чрезвычайно резки. Из-за того что воды Берингова моря расслоены слабо, в результате вертикального перемешивания в эвфотическую зону поступает огромное количество питательных веществ.
Поэтому мы обнаруживаем здесь крупные скопления нескольких видов рыб: например, палтуса, минтая и камбалы. В Атлантике Северное и Гренландское моря и море Лабрадор богаты также треской и сельдью. В этих широтах водится мало видов рыб, но их популяции имеют огромную биомассу — условия, идеальные для промыслового рыболовства. В противоположность этому экосистему тропического океана мы должны были бы описать как систему, в которой физические условия круглый год весьма однообразны, жизненные циклы организмов короче, энергия передается через пищевые сети быстро, — следовательно, наличная биомасса меньше и промысловое рыболовство ограничено.
Мы можем продвинуться в своих рассуждениях еще на один шаг. У арктических копепод развился продолжительный жизненный цикл для того, чтобы «приурочить» периоды размножения к коротким сезонам «цветения» растений, но, кроме того, им должно быть выгодно увеличить количество икры и личинок, производимых за один период размножения. Тенденция арктических копепод и бокоплавов увеличиваться в размерах свидетельствует о том, что это им необходимо, поскольку число икринок, производимых особью, экспоненциально возрастает с увеличением ее размеров. Размеры взрослой особи антарктического вида Euphausia superba достигают примерно 5 см, хотя это все еще в основном растительноядное животное. Отсюда следует, что число видов в развивающемся арктическом сообществе должно оставаться низким из-за сильной конкуренции среди личинок разных видов при ограниченных ресурсах растительной пищи.
Низкотемпературные воды. Мы знаем, что активность тепловодных организмов замедляется, когда они попадают в более холодную воду. Но было бы ошибкой переносить эту тенденцию на животных, обитающих в полярных океанах с температурой, близкой к точке замерзания воды. Для рыбы так же важно обладать способностью «двигаться быстро», чтобы схватить жертву, в холодной воде, как и в теплой. Развившиеся в процессе эволюции преимущества быстроты столь важны для выживания видов, что и в полярных океанах этот принцип должен быть применим. Нам известно также, что различные ферменты, используемые в процессах обмена веществ, действуют в разной степени, так что организм, развившийся в пределах арктической экосистемы, может иметь такую же скорость обмена веществ, что и организм, развивавшийся в низких широтах, просто в результате выбора фермента. Короче говоря, суровость арктического климата не позволяет объяснить ни скудное видообразование, ни низкую первичную продукцию.
Но суровые арктические условия, вероятно, влияют на структуру сообщества организмов в Северном Ледовитом океане. Среди хищников, находящихся на верхних уровнях пищевой цепи, многие виды — теплокровные животные: тюлени, выдры, киты, птицы и медведи. То, что организмы, способные регулировать температуру тела, настолько преобладают в полярных океанах, удивительно само по себе; подробнее мы расскажем об этом позднее.
«Возраст» экосистемы Северного Ледовитого океана. Высокая продукция Южного океана. Обширные моря Арктики мелководны, глубина Берингова пролива тоже невелика — около 50 м. Поэтому понижение уровня моря примерно на 150 м в плейстоценовый период — около 40 тыс. лет назад (рис. 4.12, г) — привело к осушению огромных площадей Арктического бассейна. Воды Северного Ледовитого океана были изолированы как от Тихого океана, так и от теплых вод Гольфстрима, которые в настоящее время переносят тепло в Баренцево море. Топографически Северный Ледовитый океан превратился в глубокий океанический бассейн практически без всякого шельфа и, подобно современному Средиземному морю, стал соединяться с Мировым океаном лишь одним узким проливом.
Такие изменения топографии и циркуляции вместе с изменением температуры должны были оказать сильное влияние на структуру сообществ живых организмов в Северном Ледовитом океане. На этом основано мнение о том, что слабое видообразование современного Ледовитого океана отражает его экологическую «молодость», т. е. здесь просто не прошло достаточно времени для того, чтобы в экосистеме развились и заполнились потенциальные «экологические ниши». Сторонники этой гипотезы указывают на большее разнообразие видов в антарктических морях, поскольку уже при 600-м глубине понижение уровня моря всего на 150 м должно было слабо сказаться на условиях жизни на шельфах Антарктиды, следовательно, здесь воздействие на сообщество живых организмов, вероятно, было незначительным. Этот вопрос еще изучается.
Проблема отбора проб. Некоторые ученые считают, что современное, кажущееся скудным видообразование во флоре и фауне Северного Ледовитого океана главным образом является результатом недостаточно полного опробования. Чем больше исследований проводится, тем большее число видов становится известно, и очевидно, что степень опробования сообществ Северного Ледовитого океана гораздо меньше, чем, скажем, сообществ Северной Атлантики между Вудс-Холом, шт. Массачусетс, и Бермудскими островами. Следовательно, вопрос остается открытым и может быть разрешен только по прошествии времени. Каким бы ни был результат, несомненно, что экосистема Северного Ледовитого океана — единственная в своем роде.
Высокая продукция Южного океана.Как центральная часть Северного Ледовитого океана отличается своей низкой продукцией, так Южный океан знаменит своей очень высокой продукцией. Особого упоминания заслуживают две особенности экосистемы Южного океана: его своеобразные трофические цепи и обеспечивающие их уникальные физические условия. На рис. 20.10 показаны в упрощенном виде главные звенья пищевых цепей.
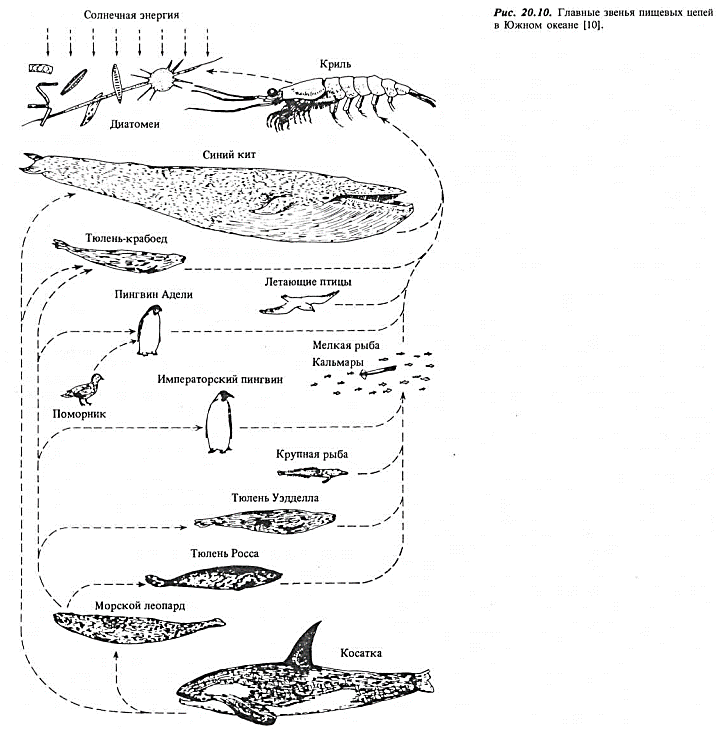
Заметим, что число трофических уровней между первичными продуцентами и хищником, стоящим на верхнем уровне, невелико: всего два для синего кита, тюленя-крабоеда, пингвина Адели и большинства летающих птиц. Другие тюлени, императорские пингвины и крупные рыбы находятся только на три уровня выше первичных продуцентов. В значительной степени принятая в руководствах классификация Южного океана как региона, «богатого морскими ресурсами», исходит из того, что короткие прямые пищевые цепи позволяют экосистеме накапливать огромную биомассу на высоких трофических уровнях. Поскольку животные этих уровней имеют также высокую коммерческую стоимость и благодаря своим размерам хорошо видны, богатство фауны здесь более явное, чем в других океанах.
Первичная продукция. Современные оценки величины растительной продукции Южного океана сильно не отличаются от оценок для других океанов. Были опубликованы различные значения, но важно иметь в виду, что все они получены по данным ограниченного опробования этого отдаленного океана. По сравнению с ним опробование первичной продукции в арктических водах было гораздо более представительным, а для антарктических вод практически не получено ни одной пробы первичной продукции в зимний период. Даются оценки порядка 100 г С/(м3-год), эквивалентные значениям из табл. 1.2 для прибрежных океанских вод в целом.
Существуют два условия, о которых следует помнить, сравнивая продуктивность Южного океана и других вод. Прежде всего, количественные измерения содержания питательных веществ на больших площадях в периоды весенних и летних максимумов продукции показывают, что нитраты обычно присутствуют в избытке, иначе говоря, в этих водах истощение питательных веществ не является лимитирующим фактором.
Сравните это с тем, что изображено на рис. 8.6, где содержание нитратов в водах Орегонского апвеллинга велико, но дальше от берега запасы нитратов практически полностью истощаются к середине лета. Именно мощный апвеллинг поддерживает в Южном океане такие высокие концентрации питательных веществ в течение всего года. Но в отличие от Орегонского апвеллинга, на широтах 55—70° в Южном полушарии света, необходимого для фотосинтеза, существенно меньше. Другое условие — выедание водорослей крилем, в результате чего новая продукция быстро удаляется.
Дата добавления: 2022-01-28; просмотров: 1106;