Проникновение вируса в клетку
Простые и сложные вирусы попадают в клетку разными способами.
Простые вирусы после адсорбции на рецепторе поверхности клетки стимулируют запуск механизмов пиноцитоза. Пиноцитоз вирусных частиц называют виропексисом (рис.8).
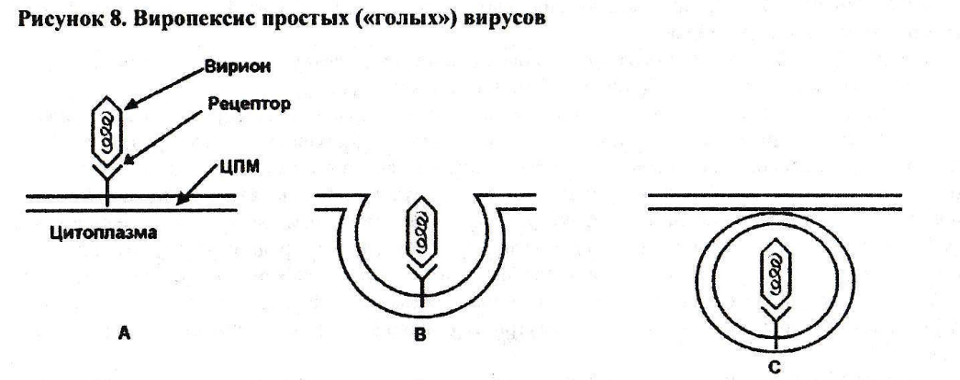
После адсорбции вириона на рецепторе клетки (рис.8А) происходит инвагинация клеточной мембраны, и вирион вместе с рецептором как бы «углубляется» в цитоплазму клетки (рис.8В). Вслед за этим участок мембраны, окружающий вирион, и цитоплазматическая мембрана в месте погружения вириона замыкаются. В результате в клетке оказывается вирион вместе с рецептором в своеобразной мембранной вакуоле (рис.8С).
Проникновение в клетку сложных вирусов происходит иначе (рис.9).
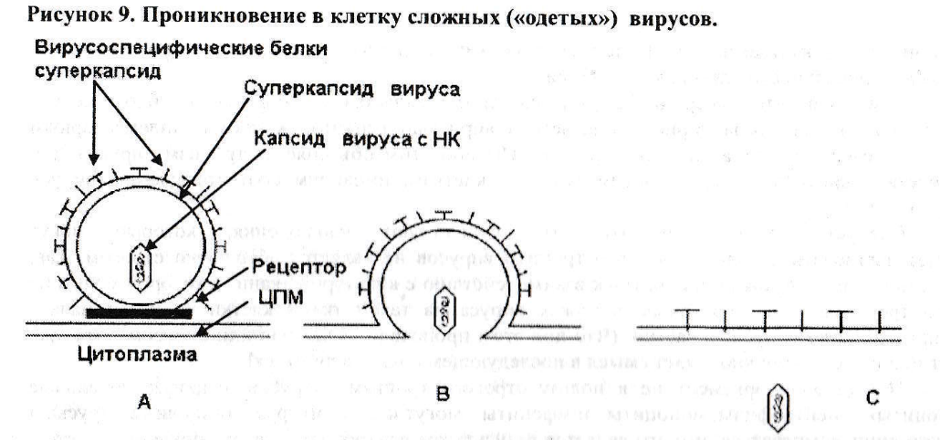
Сложные вирусы «одеты» в дополнительную оболочку - суперкапсид, формирующийся из мембраны клетки, производившей вирус. В составе суперкапсида вируса есть вирусоспецифические белки, выполняющие функцию структур, специфически соединяющихся с соответствующим рецептором клетки. После прикрепления такого вируса к рецептору (рис.9А) под действием вирусных и, вероятно, клеточных ферментов мембрана суперкапсиды конец-в-конец «сшивается» с цитоплазматической мембраной клетки (рис.9В), а в ЦПМ формируется «окно». В результате таких манипуляций капсид вируса с его нуклеиновой кислотой оказывается в цитоплазме клетки, а суперкапсид вируса - в составе ЦПМ клетки (рис.9С). Такой процесс взаимодействия сложного вируса с клеткой часто называют «механизмом слияния мембран».
Общим для обоих механизмов проникновения вирусов в клетку будет то, что в цитоплазме клетки оказываются не только вирусная НК, но и белки капсида вируса.
Два рассмотренных здесь механизма проникновения вирусов в клетку являются, видимо, самыми частыми вариантами. Вместе с тем, можно обозначить и альтернативу. Так, например, по аналогии взаимодействия фага с микробом, когда в цитоплазму клетки проникает только фаговый геном, а белки оболочки остаются снаружи клетки, можно предполагать такой вариант и для вирусов. Иногда для характеристики такого механизма используют очень емкий профессионализм- «стриптиз на поверхности». В этом случае предполагают, что контакт простого вируса с рецептором клетки приводит к изменению конформации капсомеров, они становятся доступными для действия клеточных ферментов. В результате капсид вируса разрушается на поверхности клетки, а внутрь его проникает только вирусная нуклеиновая кислота.
Таким образом, вирусная генетическая информация тем или иным способом поступает в цитоплазму клетки, что является главным условием репродукции вирусов.
Взаимодействие вирусов с клетками. Взаимодействие вирусов с клетками происходит по тем же законам, что и бактериофага с бактериями. Однако есть и существенные отличия. Они обусловлены тем, что оболочка бактерий из-за наличия клеточной стенки является жесткой структурой, не позволяющей бактериям осуществлять фагоцитирующие действия. В связи с этим при встрече бактериофага с микробом фаг не остается «пассивным» и осуществляет какие-то действия (например, сокращение футляра отростка).
Оболочка организмов животных представлена цитоплазматической мембраной, позволяющей осуществлять фаго - и пиноцитическое действие. Поэтому при встрече вируса с такой клеткой вирион остается абсолютно пассивным, и все последующие за адсорбцией вируса акции являются, главным образом, результатом активности клетки (входящие в состав вируса ферменты могут принимать участие в этих процессах, но только как «пассивные» элементы, просто столкнувшиеся с подходящим объектом).
Встреча вируса с клеткой и адсорбции его на клетке. Вирионы, не обладая соответствующими органоидами и энергетическими системами, являются пассивными элементами и не способны к самостоятельному и, тем более, направленному движению. Как субстрат броуновского движения вирионы случайно сталкиваются с клетками, и лишь немногие из этих столкновений приводят к присоединению вириона к поверхности клетки, к адсорбции.
Считается, что адсорбция вируса на клетке является специфическим процессом и происходит тогда, когда вирион попадает на вирусный рецептор клетки комплементарным участком своего капсида или суперкапсида. Именно этим объясняется тропизм вирусов, т.е. «сродство» вирусов к определенным тканям, клеткам, имеющим соответствующие вирусу рецепторы.
Следует, однако, заметить, что существуют наблюдения, которые могут свидетельствовать в пользу того, что тропизм вирусов не является абсолютно строгим. Так, например, вирус «можно приучить» к взаимодействию с культурой ткани, к которой у него не было тропизма. При повторных пассажах вируса на такой ткани клетки ее могут начать воспроизводство вирусных частиц. (Что при этом происходит с тканью или вирусом - это уже другой вопрос. Возможно, будет смысл в последующем к нему вернуться).
И еще один аргумент не в пользу строгого тропизма вирусов. «Профессиональные фагоциты» - нейтрофилы, моноциты, лимфоциты - могут и фагоцитируют различные вирусы, и можно лишь сомневаться, что это является результатом пантропизма фагоцитирующих клеток. Вполне возможно, что в этом случае работают другие механизмы взаимодействия.
Дата добавления: 2022-01-31; просмотров: 776;