Термодинамические и биохимические основы биоэнергетики
Для оценки и истолкования результатов биотермо-динамических измерений необходимо использовать понятия изменения энтальпии (ΔН), изменения энтропии (ΔS), а также изменения свободной энергии Гиббса (ΔG). Подробный теоретический анализ этих термодинамических величин содержится в ряде руководств (см., например, Э. Кальве, У. Прат, 1963), и тем не менее мы считаем своим долгом для удобства читателей перечислить основные термодинамические характеристики, использующиеся при анализе результатов биоэнергетических исследований.
Изменение энтальпии (ΔН) определяется как:
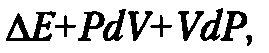
где ΔЕ — изменение полной внутренней энергии системы, dV и dP — приращение соответственно объема системы при постоянном давлении и давления при постоянном объеме.
Объем камеры для измерений основного обмена и давление внутри нее постоянны, поэтому ΔН=ΔЕ. При всяком самопроизвольном процессе происходит перенос энергии в системе таким образом, что ее способность к дальнейшему превращению (совершению работы) стремится к минимуму, т. е. система стремится к состоянию равновесия. Мерой приближения системы к равновесному состоянию служит энтропия — S. Это совокупность всех тепловых потерь системы в данном температурном интервале в расчете на градус температуры (Т):
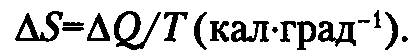
Произведение TΔS имеет размерность энергии и сопоставимо с ΔН и ΔЕ.
Все системы стремятся к минимальному теплосодержанию, поэтому состояние системы тем устойчивее, чем более экзотермична реакция. С другой стороны, все системы стремятся к максимуму энтропии, т. е. наиболее вероятному состоянию, соответствующему минимальной упорядоченности. Общий критерий самопроизвольности процесса требует соединения ΔН и TΔS. Таким критерием в термохимии служит функция Гиббса (G).
При постоянных давлении и объеме:
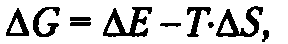
ΔG — это та часть внутренней энергии системы, которая не «энтропизована», т.е. годна для совершения полезной работы. В условиях калориметра приближение к равновесию означает, что система стремится к состоянию, исключающему обмен свободной энергией (G стремится к нулю). Тогда ΔG0) реакции, но последние лишь при условии:
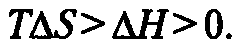
Живые организмы существуют в стационарных, удалённых от термодинамического равновесия динамических состояниях, за счёт постоянной утилизации химической энергии пищевых субстратов для совершения полезной работы. Они поддерживают минимальную скорость нарастания собственной энтропии (И. Р. Пригожин). Тем не менее, организм не стоит за рамками закона сохранения энергии, а его работа имеет коэффициент полезного действия, далёкий от 100%.
Следовательно, интегральную характеристику энергетического метаболизма— основной обмен — можно рассматривать с точки зрения вышеизложенных термодинамических закономерностей.
Для понимания того, как формируются энергозатраты основного обмена, необходимо вспомнить ряд положений физиологической химии и биоэнергетики (рис. 1).
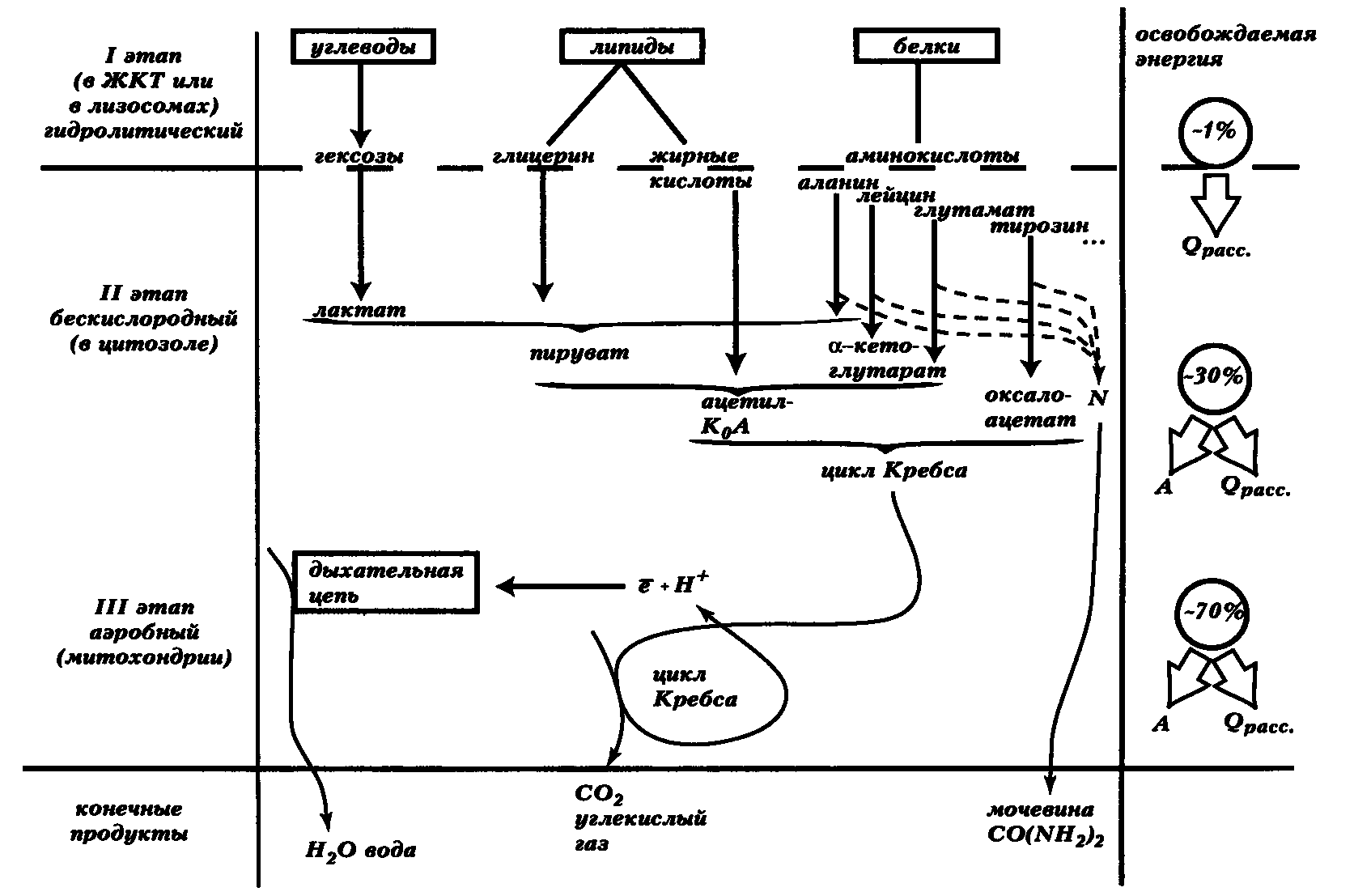
Рис. 1. Основные этапы катаболизма и их сравнительная энергетическая эффективность. Сокращения: Qрасс — первично и вторично рассеянное тепло, А — полезная работа, КоА — коэнзим А
Организм получает энергию в результате катаболизма пищевых веществ, в нормальных условиях — в основном углеводов и липидов. И те, и другие могут запасаться, служить для энергетической подпитки между приемами пищи и свободно переходить друг в друга, причем липиды запасаются долговременно. В обычных условиях 99 % потребляемых углеводов расходуется на производство свободной энергии (А. Гайтон, 1989). Белки не могут быть синтезированы только исходя из липидов и углеводов, так как содержат азот и незаменимые, то есть не вырабатываемые заново аминокислоты. Кроме того, нет специального резервного пула белков, предназначенных для энергетических целей.
Поэтому, хотя отдельные глюкогенные и кетогенные аминокислоты могут после поступления в организм превращаться в компоненты углеводов и липидов при равновесном процессе дезаминирования, за исключением этой части циркулирующего пула аминокислот, белок и его компоненты не используются нормальным организмом в качестве топлива, а идут на обеспечение синтеза азотсодержащих продуктов. Массированное использование белка в энергетических целях возможно лишь в крайне вынужденных ситуациях (например, при стрессе (см. т. 1) и голодании — см. ниже). Это заставляет вспомнить известную фразу Д. И. Менделеева, сказанную касательно утилизации нефти как топлива: «Кто же топит печку ассигнациями!»
Катаболизм — трехэтапный процесс, каждый из этапов приводит к освобождению определенной энергии, которая частично рассеивается в виде тепла сразу, не участвуя в совершении работы. Данная компонента носит название первично рассеянного тепла и непосредственно идет на нагрев тела, а, следовательно, сразу входит в тепловые потери организма, в том числе и при определении основного обмена.
Первый гидролитический этап катаболизма проходит в ЖКТ и лизосомах без участия кислорода и освобождает до 1 % всей энергии субстратов. Эта энергия полностью входит в первично рассеянное тепло и не запасается. Второй этап — бескислородного цитоплазматического расщепления — представлен в клетках гликолизом и аналогичными процессами распада липидов, приводящими к получению универсального катаболита — активного двухуглеродного фрагмента ацетил-коэнзима А (Ф.А. Липман, 1953). Это приводит к освобождению не менее 30% всего теплосодержания субстратов, при этом запасается около 43 %, а остальная часть тоже первично рассеивается (56000 калорий содержится в моле глюкозы и 2 моля АТФ прибавляется при ее распаде до пирувата, что соответствует 24000 калорий). В анаэробных условиях практически единственным источником энергии для организма служат углеводы, тогда как в аэробных — спектр субстратов расширяется.
Все клетки организма, за исключением эмбриональных и злокачественных, а также эритроцитов, в которых почти нет митохондрий, в той или иной мере проявляют способность подавлять анаэробный распад глюкозы и активировать аэробное митохондриальное окисление в условиях доступности кислорода при исправных митохондриях (эффект Л. Пастера).
Продукты второго этапа катаболизма, вовлекаясь в митохондриальное окисление, освобождают до 70 % всей своей химической энергии и распадаются до конечных метаболитов — для углеводов и простых липидов это будут С02 и Н20 (Х.А. Кребс, 1937). Закономерности окислительного этапа катаболизма были раскрыты благодаря изучению О. Варбургом (1931) и А. Сент-Дьердьи (1937) ансамбля дыхательных ферментов и описанию А. Клодом и А. Ленинджером (1948) строения и функции митохондрий. Наконец, решающую роль сыграло обнаружение АТФ, окислительного фосфорилирования (В.А. Белицер, В.А. Энгельгардт, 1930-1939) и хемиосмотического механизма последнего процесса.
Дата добавления: 2022-01-31; просмотров: 857;