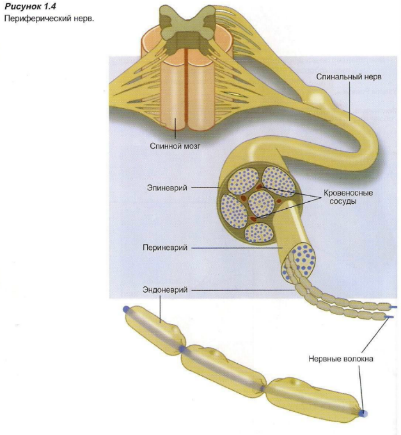
Строение периферического нерва
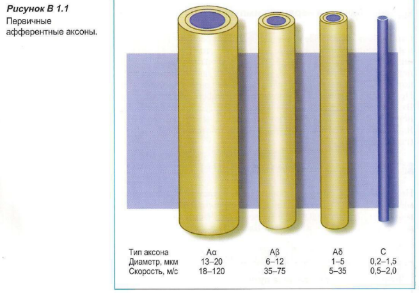
Ноцицептивные стимулы проводятся по чувствительным тонко миелинизированным Aδ волокнам и немиелинизированным С волокнам. Другие виды нервных волокон участвуют в передаче других сенсорных стимулов (см. Блок 1.1).
Блок 1.1 Ноцицептивные пути. В организме ноцицептивные стимулы, полученные ноцицепторами, затем распространяются по Aδ и С волокнам. Первые — тонко миелинизированные волокна с быстрой передачей импульса, тогда как вторые — немиелинизированные волокна с медленной передачей.
Волокна С проводят импульсы, полученные от воздействия температуры, боли или зуда. Аα волокна проводят двигательные импульсы от изменения положения и движения тела; Aß волокна передают импульсы, образованные прикосновением, а также сигналы от кожных механорецепторов; Aδ волокна проводят болевые импульсы, температурные сигналы и сигналы для поддержания мышечного тонуса.
Клеточные тела этих первичных нейронов локализуются в ганглиях дорзальных корешков. Их аксоны проходят по тракту Лиссора (Lissauer) к дорзальным рогам спинного мозга, где соединяются со вторым чувствительным нейроном в пластинке Рекседа (Rexed). Второй чувствительный нейрон переходит через срединную линию на противоположную сторону и поднимается вверх, образуя спиноталамический тракт. Спиноталамический тракт образует синапсы с ядрами таламуса, откуда ноцицептивный путь продолжается к соматосенсорной коре. Нисходящие пути, идущие из соматосенсорной коры, оказывают модулирующее воздействие на ноцицептивную систему. Эти волокна высвобождают нейромедиаторы серотонин и норадреналин.
Периферический нерв образован нервными волокнами группы нейронов, окруженных соединительнотканной оболочкой. Отдельные волокна могут быть окружены или не окружены изолирующим миелиновым слоем — шванновской оболочкой. Тело клетки является метаболическим центром нейрона (Рисунок 1.3), где синтезируется большинство клеточных органелл.
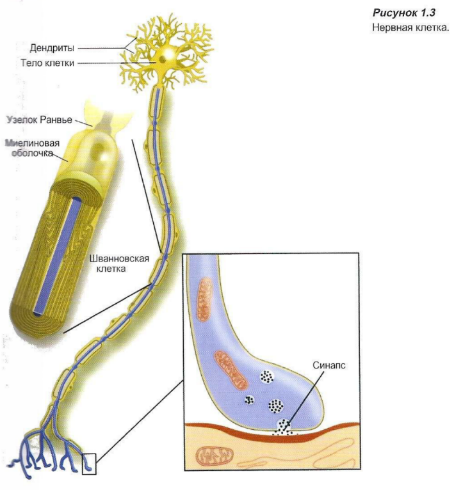
Дендриты проводят импульсы к телам клеток, а аксоны передают сигналы от тел клеток дальше. Некоторые аксоны окружены миелиновой оболочкой, другие — нет. Аксоны и дендриты удлиняются и образуют нервные волокна. Дендриты оканчиваются рецепторами, которые способны воспринимать сигналы. Аксоны оканчиваются синапсами, через которые импульсы передаются другой нервной клетке или клетке концевого органа.
Нервы — это пучки нервных волокон, удерживаемых вместе соединительной тканью (Рисунок 1.4). Каждый отдельный аксон окружен соединительной тканью (эндоневрием). Связка нервных волокон образует пучок, который также удерживается вместе соединительной тканью (периневрием). Определенное количество пучков удерживается вместе опять же соединительной тканью (эпиневрием), образуя нерв.
Формирование импульса. Возникновение и проведение импульсов в нервных волокнах — сложный процесс. Для того чтобы вызвать электрические импульсы, должно произойти изменение электрического заряда. Клетки окружены полупроницаемой мембраной, которая пропускает только воду. Селективный ионный насос активно закачивает ионы калия внутрь клетки, а ионы натрия откачивает из клетки. Это приводит к возникновению разницы концентрации ионов натрия и калия по разные стороны мембраны.
Цитоплазма клетки содержит отрицательно заряженные белки в большой концентрации, которые придают клетке отрицательный заряд (относительно окружающей среды). Вне клетки также есть отрицательно заряженные ионы, в основном, хлорид-ионы. По обе стороны мембраны электрический заряд уравновешивается положительно заряженными ионами (натрия, калия, кальция). Вследствие того, что концентрация анионов внутри клетки несколько выше, чем снаружи, количество катионов внутри клетки будет больше, чем снаружи. Это является причиной трансмембранной разницы потенциалов, составляющей -60 мВ, — так называемого потенциала покоя.
Мембрана имеет ионные каналы, которые находятся в открытом или закрытом состоянии (Рисунок 1.5).
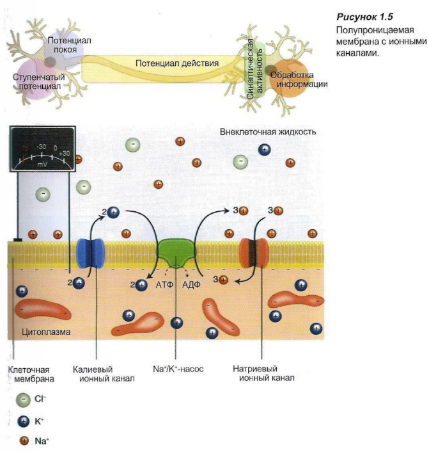
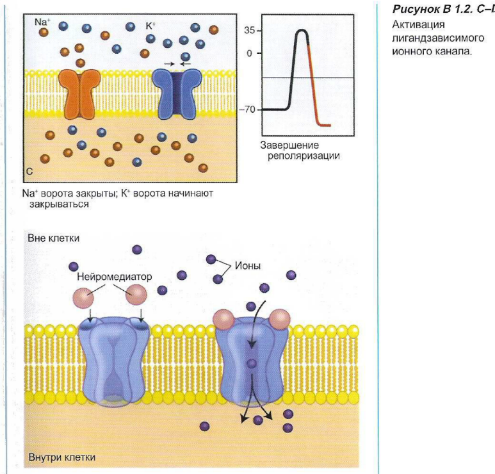
Эти каналы могут активизироваться под влиянием электрических («потенциалзависимых») или химических («лигандзависимых») стимулов (см. Блок 1.2). В том случае, когда ионные каналы открыты, ионы перемещаются по градиенту концентрации. В покое, в основном, открыты калиевые каналы, поэтому ионы калия пытаются покинуть клетку. Однако относительная «перегрузка» клетки анионами (белками) препятствует утечке катионов. Когда открываются натриевые каналы мембраны, ионы натрия начинают входить в клетку: иначе говоря, в мембране появляется «дыра».
Блок 1.2. Ионные каналы чрезвычайно важны для образования, проведения и передачи нервных импульсов. Активация этих рецепторов может происходить при помощи электрических стимулов (потенциалзависимые каналы) и нейромедиаторов (лигандзависимые каналы). После того как активированные каналы открываются, это позволяет перемещаться ионам, вызывая деполяризацию клеточной мембраны.
Потенциалзависимые ионные каналы, в том числе быстрые натриевые каналы и кальциевые каналы, участвуют в образовании импульсов в сердце и проведении импульсов в нервных волокнах. Примерами лигандзависимых ионных каналов являются ацетилхолиновые, глутаминовые и ГАМК рецепторы.
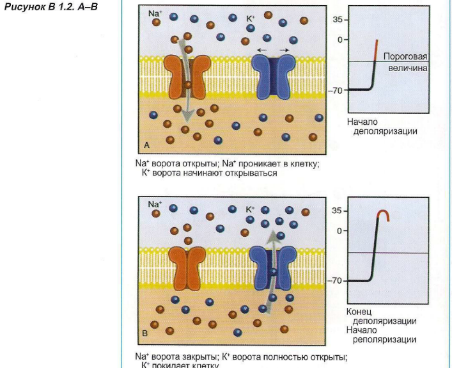
Приток ионов натрия нарушает электрическое равновесие, возникает локальная деполяризация, и ионы калия получают возможность покинуть клетку. Это восстанавливает баланс между анионами и катионами (реполяризация). Во время деполяризации и в начале реполяризации новая деполяризация возникнуть не может (рефрактерный период).
Если локальная деполяризация незначительна, равновесие быстро восстанавливается (Рисунок 1.6). И только когда локальная деполяризация достигает определенной пороговой величины (примерно -50 мВ), возникает потенциал действия. Таким образом проявляется эффект «все или ничего».
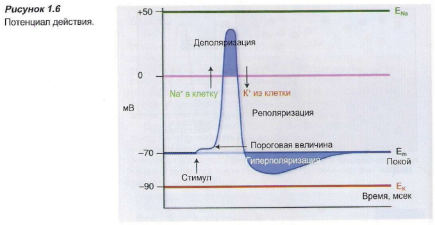
Пороговая величина, необходимая для развития потенциала действия, определяется несколькими факторами, такими как продолжительность и сила деполяризующего стимула и состояние рецептора. Только посредством достижения пороговой величины потенциалзависимые натриевые каналы открываются, возникает приток натрия, и мембрана меняет полярность.
Натриевые каналы остаются открытыми только в течение приблизительно 1 мсек, после чего они вновь закрываются. Калиевые каналы затем еще остаются открытыми, и отток калия через потенциалзависимые калиевые каналы восстанавливает электрическое равновесие, при этом даже возникает гиперполяризация. После этого потенциалзависимые калиевые каналы закрываются, и натрий-калиевый насос восстанавливает первоначальное положение. Количество ионов натрия и калия, которые перемещаются для того, чтобы вызвать потенциал действия, исключительно небольшое.
Дата добавления: 2022-01-27; просмотров: 853;